

 English
English文献解读|Nat Med(50):抗PD-1治疗后,与HIV病毒库相关的先天性抗病毒和免疫功能会减弱
✦ +
+
论文ID
原名:Innate antiviral and immune functions associated with the HIV reservoir decay after anti-PD-1 therapy
译名:抗PD-1治疗后,与HIV病毒库相关的先天性抗病毒和免疫功能会减弱
期刊:Nature Medicine
影响因子:50
发表时间:2026.02.12
DOI号:10.1038/s41591-025-04139-y
背 景
抗逆转录病毒疗法(ART)改善了全球HIV感染者的死亡率和预后。ART可恢复CD4+ T细胞,使CD4:CD8 T细胞比例正常化,并促进免疫稳态。然而,ART并非治愈性疗法,中断治疗会导致血浆病毒载量迅速反弹。此外,HIV持续存在会驱动免疫激活和慢性炎症,并增加非艾滋病合并症(如癌症和心血管疾病)的风险。研究表明,即使长期接受ART治疗,潜伏的HIV仍存在于少量CD4+ T细胞中,这构成治愈的主要障碍。该HIV病毒库富含表达免疫检查点蛋白(PD-1、CTLA-4、TIGIT和TIM-3)的记忆性CD4+ T细胞;这些蛋白质的表达会损害维持储存库持久性的稳态增殖。ART可以抑制 HIV 病毒,但无法清除潜伏病毒库,该病毒库存在于表达程序性死亡蛋白 1 (PD-1) 的 CD4+T 细胞中。抗 PD-1 疗法已降低了 HIV 感染者 (PLWH) 合并癌症患者体内的 HIV 病毒库;然而,哪些患者能够从中获益以及病毒库降低的机制尚不明确。
实验设计
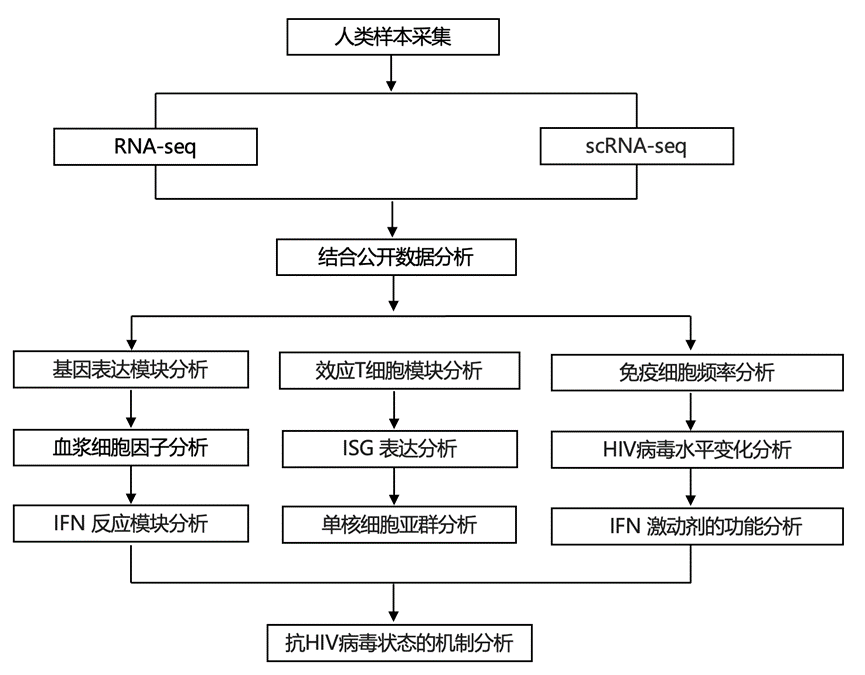
结 果
01

多组学分析显示,抗PD-1治疗后24小时内即可快速诱导先天性IFN反应
为了阐明阻断PD-1结合所触发的导致潜伏性HIV病毒库减少的机制,研究团队分析了癌症免疫治疗试验网络-12(CITN-12;NCT02595866)试验期间收集的纵向样本。研究参与者的详细基线人口统计学和临床特征,包括年龄、性别、癌症类型和抗逆转录病毒治疗方案。该试验是一项多中心I期研究,其中对患有癌症的HIV感染者(PLWH)使用了抗PD-1抗体帕博利珠单抗(图1a)。本研究呈现的CITN-12免疫学分析是该临床试验预先设定的探索性部分,其检测方法和样本采集时间点均经过前瞻性定义,旨在捕捉抗PD-1给药后的免疫和病毒学动态变化。共有30名病毒载量受到抑制的HIV感染者(PLWH)接受了标准帕博利珠单抗方案治疗(每3周静脉注射200 mg,疗程最长2年),所有参与者均完成了至少两个疗程(图1a-b)。根据基线CD4+ T细胞计数,参与者分为三个队列:队列1(100-199个CD4+ T细胞/μL,n =6)、队列2(200-350个CD4 + T细胞/μL,n =12)和队列3(>350个CD4 +T细胞/μL,n =12)(图1a-b)。这些参与者共患有11种不同的癌症,包括艾滋病相关癌症,例如卡波西肉瘤和非霍奇金淋巴瘤(图1a-b)。癌症亚型的分布与基于CD4+ T细胞计数的队列无显著相关性。值得注意的是,一名参与者达到完全缓解,四名参与者达到部分缓解(图1b)。他们从来自三个CD4+ T细胞计数队列的参与者中采集了样本,这些参与者患有11种不同的癌症类型,样本采集时间点分别为治疗前(C01D01)、治疗后24小时(C01D02)、治疗后1周(C01D08)和治疗结束时(EOT)。他们采用了一种综合的多组学方法。其中包括对血浆细胞因子和外周血单核细胞 (PBMC) 转录组进行分析,以确定治疗后 24 小时内与先前报道的潜伏 HIV(细胞相关 HIV DNA)下降和病毒再激活(HIV RNA)短暂增加相关的免疫改变。随后,他们研究了 (1) 这些转录组特征在治疗结束前的持续性,(2) 与 HIV 结局(例如,血浆 HIV RNA 和细胞相关 HIV DNA 水平)的关联,以及 (3) 使用单细胞和体外验证方法,研究它们在免疫细胞亚群中的普遍性和机制作用。
他们采用转录组分析(RNA-seq)来鉴定C01D01和C01D02之间发生改变的基因表达模块(图S1a-b)。为了获得这些模块,首先使用来自分子特征数据库(MSigDB)C7集合的免疫学特征基因集进行基因集富集分析(GSEA)。然后,基于前沿基因的重叠,将C01D02相对于C01D01显著改变的通路合并,并使用合并后的前沿基因来定义每个转录组模块。同时,使用GSEA评估了ChIP-X富集分析(ChEA)数据库中整理的转录因子靶标集的富集情况。该数据库汇总了来自多个染色质免疫沉淀测序(ChIP-seq)和染色质免疫沉淀芯片(ChIP-chip)研究的经实验验证的转录因子-靶标相互作用。接下来,他们分析了样本水平的模块和转录因子靶标评分,以定量每位参与者32的这些基因集的相对表达量(图1c)。他们观察到,单核细胞功能(模块4和5)、IFN/病毒反应(模块2和3)以及CD8 T细胞活化(模块1)模块在C01D02时显著上调(图1c)。他们还观察到调节IFN反应(IRF、STAT1)、单核细胞效应功能(RELA、STAT4)和T细胞活化(STAT5)的转录因子增加,而免疫检查点阻断剂(如PDCD1、TIGIT、IL10RA)在C01D02时表达降低(图S1c-d)。在 C01D02 时,T 细胞干性、WNT/β-catenin 信号传导和调节性 T (Treg) 模块以及受 TCF1 和 β-catenin 调控的基因和表观遗传调控因子(如 EGR1和 EZH2)的表达均降低。
血浆细胞因子谱分析显示,在C01D02和C01D08时,包括IFNβ、IL-6和IFNγ(TH1反应)在内的固有免疫细胞因子显著上调,而TGFβ1水平在C01D02时显著降低。值得注意的是,TH 2(IL-4)和TH17(IL-17A)反应细胞因子的水平仅在C01D08时升高。接下来,他们构建了一个整合的转录组-细胞因子网络(图1d),以证明包含单核细胞功能、ISG和效应CD8+ T细胞相关基因集的转录组模块与血浆IFNγ、IL-6和IFNβ水平之间存在正相关性。相反,血浆TGFβ水平降低与WNT信号通路和Treg模块表达减少相关。这些结果表明,抗PD-1疗法可降低血浆TGFβ水平,并促进促炎和抗病毒先天免疫反应在24小时内快速增强。
上述抗PD-1诱导的转录组模块在14名HIV感染者(PLWH)的治疗结束时(EOT)保持稳定。然而,这些模块在EOT时将参与者分为两个不同的组(图1e)。第1组(ISGhi)(n = 9)的样本表达ISG/抗病毒基因(模块2和3)、单核细胞基因(模块4和5)以及CD8效应基因(模块1)。纵向研究显示,第1组参与者在四个疗程(约3-4个月)内维持了较高的ISG水平和抗病毒模块的表达,表明免疫重编程持续存在。相比之下,第 2 组(ISGlo)PLWH(n = 5),包括接受延长治疗(超过四个疗程)的患者,在治疗结束时表现出较低的 ISG 模块表达,表明这些免疫反应性的差异并非治疗持续时间所致。第 2 组样本还显示 WNT(模块 6)和 Treg 基因集(模块 7)的表达增加(图1e)。
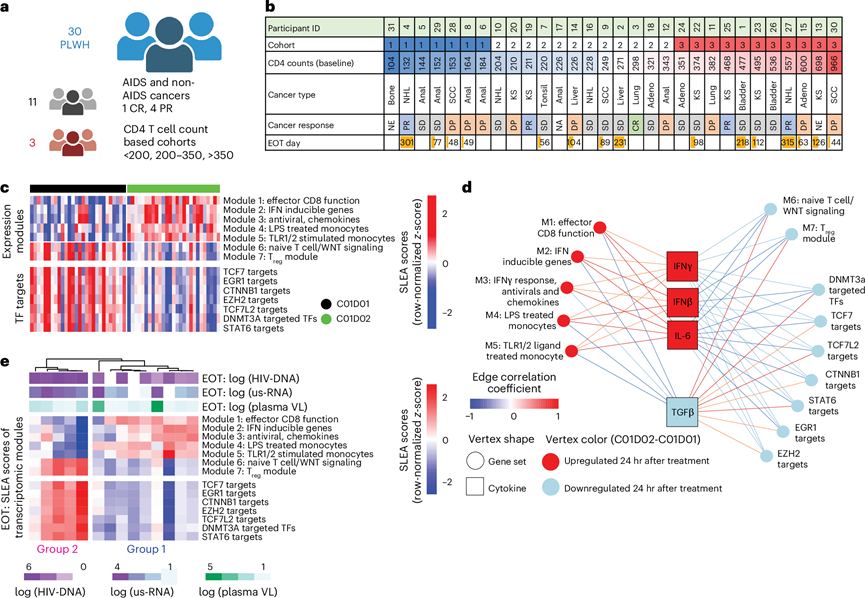
图1. 抗 PD-1 输注在治疗开始后 24 小时内引起血浆细胞因子和全血转录组模块的系统性改变。
(a) 样本采集自30名HIV感染者,涵盖11种不同的癌症类型。(b) 表格列出了参与者的研究ID、CD4 +T细胞计数组。(c) C01D01和C01D02之间转录组模块的SLEA评分显著变化。(d) 整合了转录组模块的SLEA评分和C01D01和C01D02时血浆细胞因子水平的mixOmics模型。(e) 转录组模块SLEA评分的列聚类。
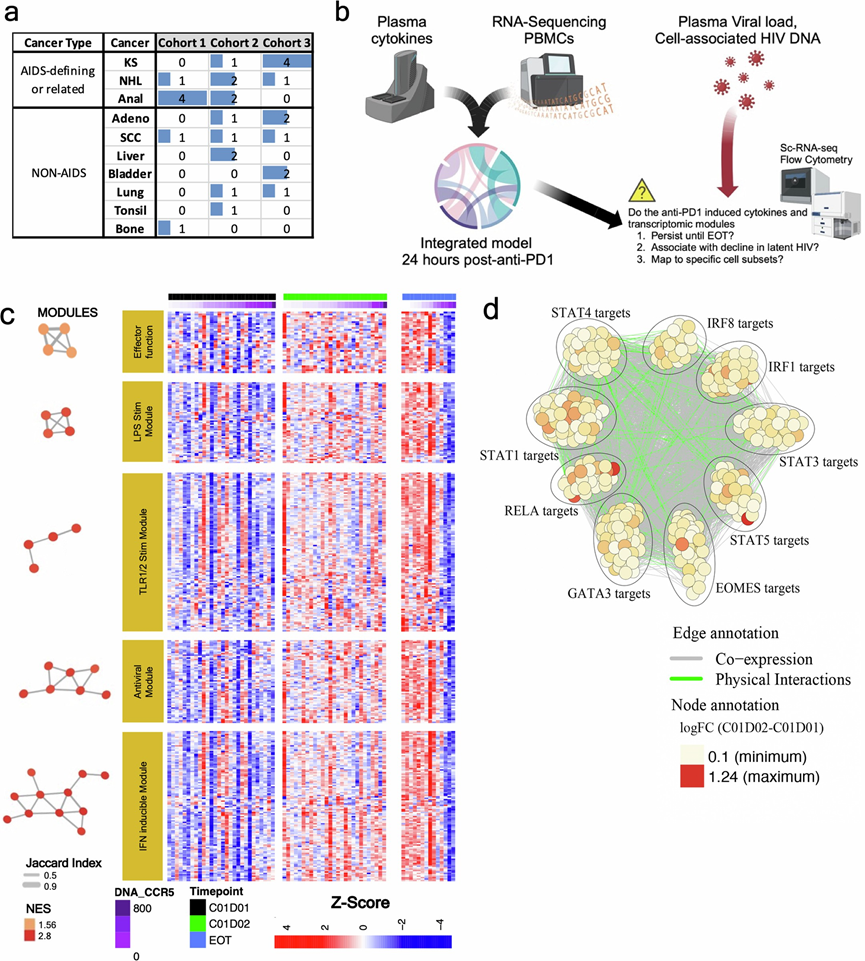
图S1. PD-1 阻断后 24 小时转录模块激活和转录因子靶向。
(a) 按艾滋病定义状态和非艾滋病定义状态分层的各队列癌症类型分布。(b) 样本处理和实验流程示意图。(c) 治疗后24小时上调的基因表达模块的热图,包括干扰素刺激基因(ISG)、抗病毒效应因子、TLR/IL-1信号通路和免疫检查点阻断模块。(d) 转录因子靶标网络突出显示了治疗后 24 小时上调靶标的富集情况。
02
抗PD-1诱导的转录组模块可根据治疗结束时HIV病毒库的结局对参与者进行分层
在每个时间点对第1组和第2组参与者的样本进行直接比较(图2a)显示,在C01D01时,第1组的ISG(模块2和3)(图2b)和单核细胞(模块4和5)(图2c)表达显著高于第2组,而第2组的WNT信号通路(模块6)表达升高(图2d)。PD-1阻断后,两组的ISG和单核细胞模块均显著升高,而WNT信号通路/Treg模块则显著降低(图2b-d)。到治疗结束时,第1组的ISG和单核细胞模块表达仍然升高(图2b-c),并且效应CD8+ T细胞功能模块(模块1)显著升高(图2d),而该模块在基线时并未升高。相比之下,第 2 组受试者的 WNT 和 Treg 模块表达持续升高,而 ISG 和单核细胞反应减弱(图2b-c)。这些数据共同表明,第 1 组(ISGhi)和第 2 组(ISGlo)受试者之间的转录差异在基线时即可检测到,并在 PD-1 阻断后更加显著,其中一部分受试者表现出持久的 ISG 和髓系抗病毒模块表达——这凸显了持续的免疫重编程,而这可能是治疗反应的基础。
与先前的数据一致,C01D02时每百万细胞的HIV DNA拷贝数显著降低,而C01D08时血浆HIV RNA水平显著升高,且与血浆中高浓度的IFNγ和低浓度的IL-4相关(图2e)。他们还观察到,第1组的9名受试者中有8名在治疗结束时CD4 + T细胞中可诱导HIV RNA的频率降低,这与病毒库整体收缩一致。相比之下,第2组的5名受试者中有4名在治疗结束时可诱导HIV-1 RNA的频率升高(图2f)。值得注意的是,每百万细胞中的HIV DNA拷贝数与携带可诱导HIV的细胞频率呈正相关。虽然这两个参数也与其他病毒学标志物相关,但在血浆HIV RNA、未剪接HIV RNA或HIV RNA/HIV DNA比值方面未观察到显著的组间差异。值得注意的是,根据 CD4+ T 细胞计数、癌症类型或对治疗的临床反应,无法区分这两个组。这些结果表明,预先存在髓系 ISG 模块的 HIV 感染者亚群,在抗 PD-1 治疗的进一步诱导下,其 HIV DNA 水平也显著下降,这反映了 HIV 病毒库的减少。
接下来,他们利用单细胞转录组(scRNA-seq)技术分析了C01D01、C01D02和C01D08时点的免疫细胞亚群(图2g)。与C01D01相比,C01D02时单核细胞(尤其是经典单核细胞亚群)的频率显著增加,这一结果通过计算机模拟反卷积得到证实(图2h)。与此同时,自然杀伤细胞和CD8 T细胞的频率则有所下降(图2h)。对各亚群表达评分进行定量分析(图2i)显示,TLR模块在单核细胞中富集,IFN 模块在 T 细胞和髓系细胞中富集,Treg/ WNT 模块在淋巴细胞亚群中富集(图2i)。ISG 和抗病毒模块在TEMRA /TEM CD8 T 细胞、TH1/TH17/Treg CD4+ T 细胞和所有单核细胞亚群中均上调,其中IFNG转录本在效应 CD8+和 CD4+ THS1细胞中富集,而IFNB和IL6转录本在单核细胞中最高。为了鉴定第 1 组和第 2 组之间基于scRNA-seq的表达谱差异,他们分析了来自两组部分参与者在 C01D01、C01D02 和 C01D08 时单核细胞和 CD8 + T 细胞的模块表达。尽管样本量有限,但第 1 组(与第 2 组相比)的单核细胞在基线和抗 PD-1 治疗后 24 小时均表现出更高的 IFN 和 TLR 模块表达,而第 1 组的 CD8+ T 细胞在治疗后表现出更强的先天免疫和抗病毒模块诱导。这些结果表明,单核细胞中的早期抗病毒程序化先于效应 T 细胞反应,并且可能在治疗前就已存在。同时,他们将这些模块映射到来自健康个体、癌症患者或感染患者的35个公开的全血单细胞数据集,并观察到一部分个体,无论其疾病状态如何,均表现出ISG模块的高表达和Treg及WNT信号通路模块的低表达(图2j)。这些发现表明,高ISG和低Treg/WNT信号模块表达代表人类更广泛的免疫激活状态,与各种疾病和干预措施相关。
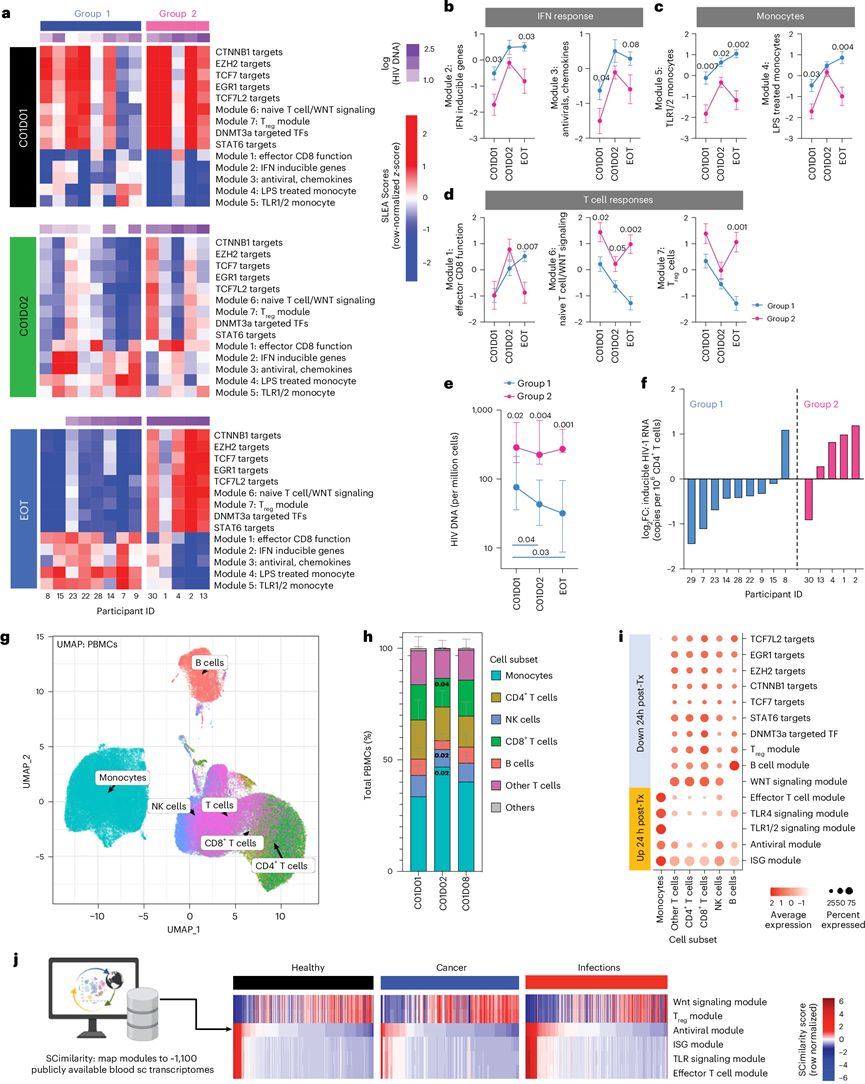
图2. 治疗前髓系和 IFN 反应模块的表达在细胞相关 HIV DNA 较低的人群中较高,并且具有普遍性。
(a) 标准化的热图显示了参与者在不同时间点(C01D01、C01D02 和 EOT)的转录组模块表达情况。(b) 在C01D01时,第1组的IFN反应和抗病毒模块评分显著高于第2组,并在治疗结束时仍保持较高水平。(c) 第1组的TLR信号通路模块评分在基线时较高,并在治疗结束时仍显著升高。(d) 效应T细胞模块评分在C01D01时无差异,但在治疗结束时第1组显著升高。(e) 在各时间点,以两两比较的方式比较各组的细胞相关HIV DNA计数。(f) 表示按组分层的受试者中诱导型HIV-1 RNA 变化。(g) scRNA-seq分析。(h) 对每位参与者在每个时间点的主要注释PBMC亚群的频率进行定量,以占PBMC总数的百分比表示。(i) 提取每个整体转录组模块中的基因,并使用Seurat的AddModuleScore函数计算每个亚群的单细胞模块得分。散点图显示了细胞频率(点的大小)和相对表达量(点的颜色)。(j) 为了评估抗PD-1治疗应答者中识别的基因特征的普遍性,使用相似性平台分析了scRNA-seq数据,并与大约1000个公共数据集进行了比较。
03
抗PD-1治疗后24小时内ISG+单核细胞中抗病毒基因表达增加与治疗结束时HIV DNA下降相关
利用基于scRNA-seq的分析,他们鉴定了单核细胞的三个不同亚群,并定量分析了每个亚群中ISG和TLR模块的表达(图3a)。抗PD-1治疗后,未观察到亚群频率的显著变化。然而,与C01D01相比,他们观察到在ISGhi中,治疗后C01D02和C01D08时, IFIT2、B2M、CD86和CXCL10等基因的表达显著升高(图3b)。该ISGhi细胞群的TLR模块基因表达也降低(图3c)。相比之下,表达TLR模块的单核细胞亚群(聚类0和1)在PD-1治疗后未显示显著的转录变化(图3c)。他们使用NicheNet技术推断潜在的配体-受体相互作用,以鉴定ISGhi单核细胞表达基因上游的假定配体(图3d)。与血浆中IFNβ和IFNγ水平升高相一致(图3c),将IFNB1和IFNG鉴定为驱动ISGhi单核细胞中基因表达上调的关键上游细胞因子(图3d)。这些结果表明,阻断PD-1结合可显著增加单核细胞频率以及特定单核细胞亚群中抗病毒ISG的表达。
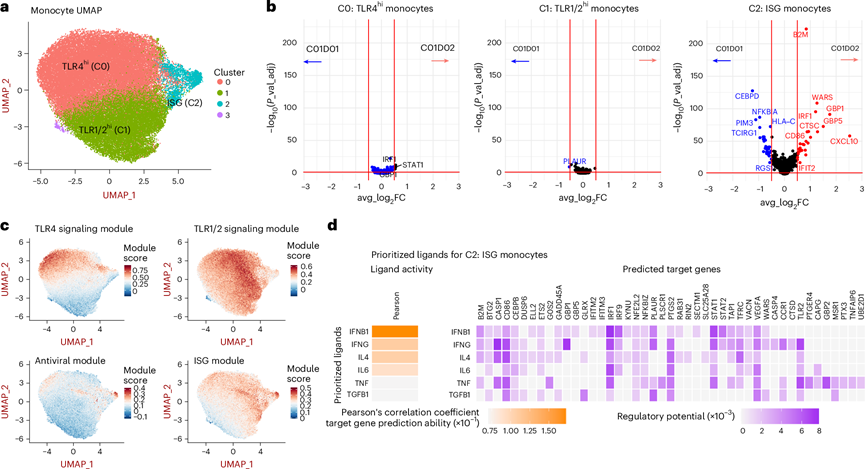
图3. 抗 PD-1 疗法在 24 小时内增加单核细胞频率和 ISG 表达。
(a) 从PBMC的scRNA-seq数据中提取标记为“单核细胞”的细胞亚群,并使用Seurat中的UMAP和无监督聚类进行分析。(b) 火山图显示,与聚类0和聚类1不同,聚类2的单核细胞在C01D02与C01D01相比,表现出更多显著上调的基因。(c) 将单核细胞的TLR、抗病毒和ISG模块评分映射到UMAP图上。(d) 使用R语言中的NicheNet包分析聚类2单核细胞(相对于所有其他单核细胞)中差异表达的基因,以识别上游调控因子。
04
抗PD-1治疗后24小时内效应ISGhi CD8 T细胞的增加与治疗结束时HIV DNA的衰减相关
鉴定出四个CD8+ T细胞亚群(图4a),其中亚群1和3表现出较高的效应基因表达(表达GZMB、PRF1、IFNG和TNF)。与亚群1相比,亚群3在C01D01时ISG富集,并在C01D02时进一步诱导(图4b-c)。亚群3细胞的频率在C01D02时显著增加,而亚群1在C01D08时扩增(图4d)。细胞亚群注释显示,这两个亚群均包含效应细胞和TEMRA细胞群的混合,流式细胞证实了效应(CD27 - CD45RA+)CD8+ T细胞在C01D02时增加(图4e)。值得注意的是,聚类1和聚类3 都表现出 ISG 表达增加,这与效应细胞分化一致。
他们通过体外用 Gag 肽池刺激PBMC进行 HIV 特异性反应分析,结果显示,在 C01D02 时,Ki-67+、产生 IFNγ 的 CD8+ T 细胞的频率增加(图4f)。为了进一步表征这些反应,他们对流式细胞数据进行了无监督聚类分析,分析对象包括 HIV 特异性(IFNγ+和/或 TNF+)CD8+ T 细胞以及从未刺激和 Gag 刺激条件下随机抽取的总 CD8+ T 细胞。在所得聚类中,聚类8(富集于第 1 组)表现出高水平的 IFNγ、T-bet、Ki-67、Id2 和 Tox,并在 C01D08 时扩增。相比之下,第 2 组参与者在整个纵向随访期间均表现出 1 型簇的显著富集(虽然IFNγ 水平高,但 Ki-67、T-bet 和 Tox 表达降低)。在第 1 天第 1 天(C01D01),第 1 组中 12.87% 的 Gag 特异性 IFNγ+CD8+ T 细胞表达 Ki-67,而第 2 组仅为 3.01%。在第 1 天第 2 天(29.9% 对 15.5%)和第 8 天(54.1% 对 36.1%),该增殖亚群在第 1 组中扩增更为显著。这些结果表明,第 1 组 PLWH 体内存在预先存在的、对 PD-1 阻断疗法有反应的、具有功能的效应 CD8+ T 细胞。
为了鉴定效应CD8+ T细胞基因的上游配体,他们使用NicheNet分析,发现IFNβ1和IFNγ是ISG和效应基因(如TNF)的主要预测诱导因子(图4g)。同时,血浆TGFβ水平降低与效应CD8+ T细胞频率增加相关(图4h);此外,还观察到参与静止期(ASXL1、BCL9L)和TGFβ信号通路(PEMPA1、SKIL)的基因表达下调(图4i)。综上所述,这些发现将抗病毒效应反应与病毒库减少联系起来,并提示PD-1阻断可快速诱导第1组参与者体内ISG+效应CD8+ T细胞的产生。
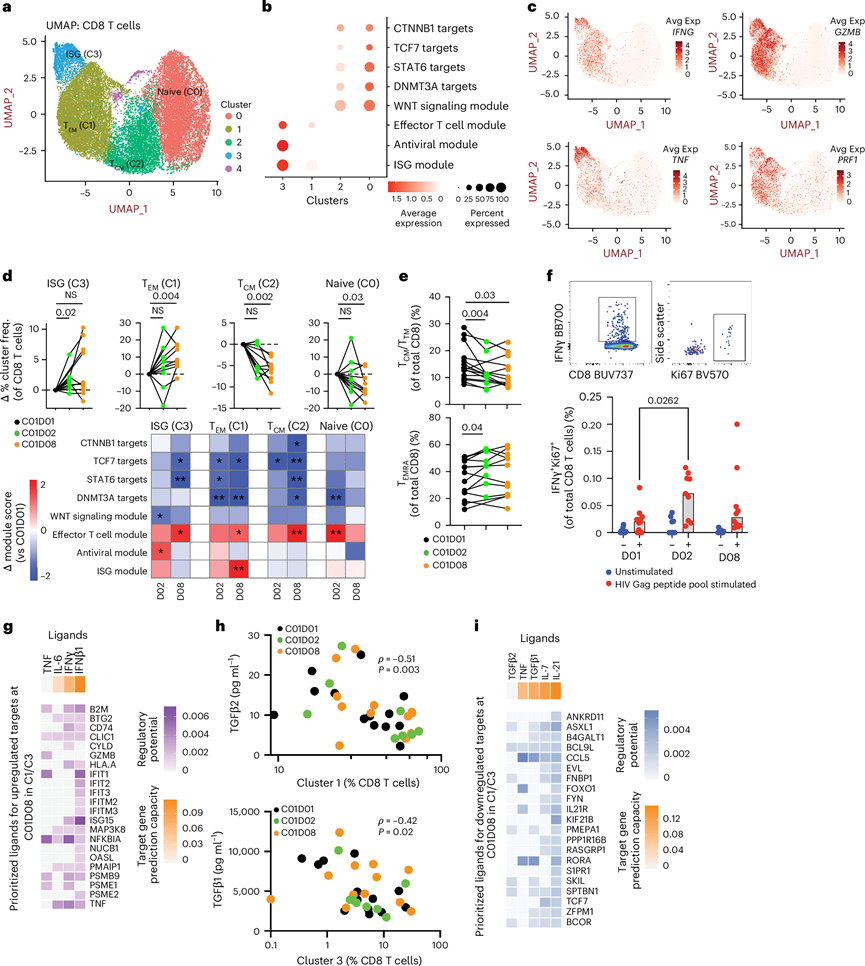
图4. 抗 PD-1 治疗 24 小时内,外周血中表达 ISG 的 CD8+ T 细胞的频率增加。
(a) 从PBMC的scRNA-seq数据中提取注释为“CD8+ T细胞”的细胞。(b) 提取每个转录组模块中的基因,并使用Seurat的AddModuleScore函数计算每个聚类的模块得分。(c) IFNG、TNF、GZMB和PRF1的特征图显示,表达这些效应基因(红色)的细胞在聚类1 和聚类3(TEM 和ISG亚群)中富集。(d) 上图显示了 C01D02 和 C01D08 时各 CD8+ T 细胞聚类频率相对于 C01D01 的相对变化。下图的热图描绘了模块评分的变化。(e-f) 流式细胞分析。(g) 效应 CD8+ T 细胞和 ISG CD8+ T 细胞聚类中上调的基因。(h) 总 CD8+ T 细胞中聚类 1(TEM)和聚类3(ISG)的频率分别与血浆 TGFβ2 和 TGFβ1 浓度呈负相关。(i) 使用 NicheNet 分析了效应 CD8+ T 细胞聚类和 ISG CD8+ T 细胞聚类(相对于其他 CD8+细胞聚类)中下调的基因。
05
持续的CD4+ T细胞ISG表达与抗PD-1反应和HIV病毒库下降相关
与在CD8+ T细胞中的发现类似,流式细胞技术和CIBERSORT分析PBMC数据均显示,在整个队列中,抗PD-1治疗后CD27+CD45RA−(中央记忆和过渡记忆)CD4+ T细胞亚群数量下降。CD4+ T细胞的注释鉴定出八个亚群,包括幼稚T细胞、Treg和效应T细胞。在治疗的第一周,效应模块(例如IL15RA和TNF)上调,而WNT信号通路下调。无偏亚聚类分析揭示了不同的细胞群:幼稚T细胞(聚类1)、ISGhi效应/记忆T细胞(聚类3)、ISGlo效应/记忆T细胞(聚类0)和调节性T细胞(聚类2)(图5a)。值得注意的是,簇3在第一周内保持稳定,而聚类0在同一时期显著下降(图5b-d),且与血浆HIV RNA水平升高相关(图5e)。使用HIV Gag肽库进行体外刺激的功能验证显示,除两名受试者外,所有受试者在治疗后24小时内HIV特异性IFNγ+ TNF+CD4+ T细胞的频率均增加(图5f)。此外,残余血浆病毒血症与HIV特异性CD4+T细胞的频率、血浆IFNγ水平以及效应模块和ISG模块的表达相关(图5f),提示存在针对HIV病毒库释放抗原的急性免疫反应。
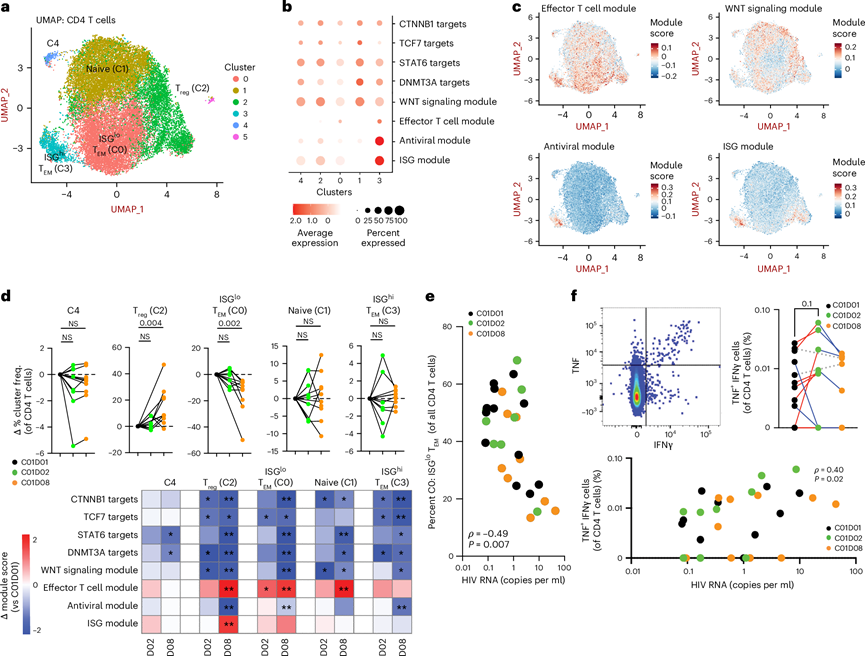
图5. 髓系细胞和 T 细胞特有的 ISG 特征在抗 PD-1 治疗后 24 小时内增加。
(a) 从PBMC的scRNA-seq数据中提取注释为“CD4 + T细胞”的细胞。 (b)提取每个转录组模块中的基因,并使用Seurat的AddModuleScore函数计算每个聚类的模块得分。(c) 特征图显示了CD4+ T细胞中效应T细胞、WNT信号通路、抗病毒和ISG模块的模块得分。(d) 上图显示了C01D02和C01D08时各CD4+ T细胞聚类频率相对于C01D01的相对变化。(e) 相关性分析表明,血浆HIV RNA水平随时间推移的升高与聚类0(ISG低表达的效应细胞)频率的降低相关。(f) 流式细胞分析。
06
亚群特异性ISG谱及其体外诱导与HIV DNA水平降低相关
为了鉴定基线(C01D01)时存在的以及在C01D02时动态诱导的分子特征和细胞因子,他们利用两个时间点的数据生成了加权特征评分(图6a)。该模型显示,较高的基线IFN诱导型、抗病毒和单核细胞TLR模块表达,结合C01D02时升高的IFNγ/IFNβ水平和降低的TGFβ水平,可预测治疗结束时较低的HIV DNA水平(图6a)。值得注意的是,基于这些加权特征预测的潜伏期HIV DNA水平与HIV DNA水平显著相关(图6b)。鉴于抗病毒和免疫调节ISG反应的重要作用,他们接下来对髓系细胞和T细胞群体中细胞亚群特异性的ISG表达谱进行了反卷积分析(图6c)。该分析揭示了两个细胞群中存在不同的ISG表达亚群。ISG +单核细胞上调了参与IFN/抗病毒信号通路(IRF7、IRF9、STAT1、BST2)、抗原呈递(B2M)和效应T细胞募集/分化(CXCL10、IL15)的基因。同时,ISG+ CD8+和ISG+CD4+ T细胞显示出髓系来源信号受体(IL15RA、TNF)和T辅助细胞极化介质(IL12RB1)表达升高。值得注意的是,这些ISG特征在HIV DNA水平较低的第1组参与者中显著富集,无论是在C01D01还是EOT(图6c)。
为了评估先天免疫刺激是否能增强抗病毒反应,他们用I/II型干扰素或TLR激动剂(poly I:C和R848)处理来自健康供体的记忆性CD4+ T细胞或去除CD8+ T细胞的PBMC。刺激导致CD4+ T细胞中抗病毒蛋白[即IFIT1(在接受抗PD-1治疗的1组HIV感染者中也升高)和APOBEC3G]的表达增加。在先天免疫细胞(例如单核细胞)存在的情况下,这种效应放大(图6d)。随后,在体外用HIV感染培养物,并添加或不添加干扰素拮抗剂。I/II型干扰素或TLR激动剂的刺激显著降低了HIV感染(图6e-f)。当存在先天免疫细胞时,TLR激动剂在抑制感染方面尤其有效。加入I型和II型干扰素拮抗剂后,尤其是在含有固有免疫细胞的培养物中,这种抗病毒控制作用消除。虽然两类干扰素均能提供抗病毒保护,但I型干扰素能更有效地诱导CD4+ T细胞中抗病毒蛋白的表达。
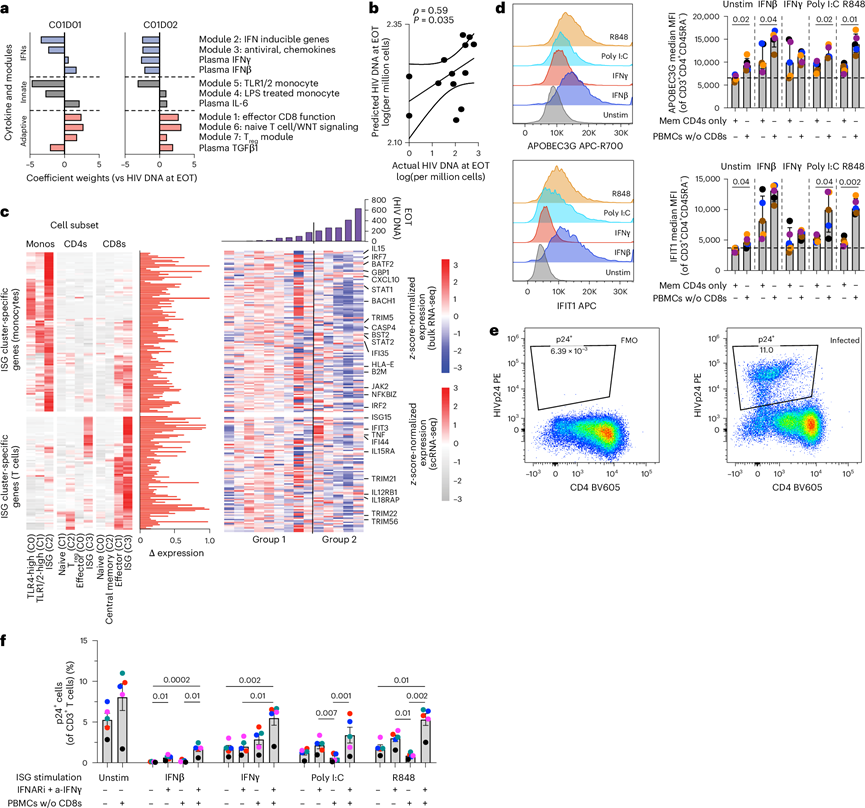
图6. 在髓系细胞存在的情况下,IFN 激动剂可保护 CD4+ T 细胞免受 HIV 攻击。
(a) 岭回归模型中与抗PD-1治疗相关的特征权重。(b) 基于岭回归模型,每百万CD4+ T细胞中预测的HIV DNA水平与实际水平之间的相关性。(c) 模型展示了细胞亚群特异性ISG表达。(d) 体外刺激试验。将来自 5 位供体(n = 5)的记忆性 CD4+ T 细胞和 CD8 耗竭的 PBMC 在有或无先天免疫刺激物(IFNα、IFNγ、poly I:C 或 R848)的情况下进行培养。(e-f) 体外HIV感染实验结果。
+ + + + + + + + + + +
结 论
本研究对多名 HIV 感染者(合并癌症),在 CITN-12 I 期临床试验中进行了一项预先设定的探索性纵向多组学分析。该试验旨在评估帕博利珠单抗的安全性和初步抗肿瘤活性。该疗法总体耐受性良好,大多数不良事件为 1-2 级,5 名受试者观察到客观抗肿瘤反应(1 例完全缓解和 4 例部分缓解)。治疗后 24 小时内,本研究观察到 HIV 特异性效应 CD8+ T 细胞增殖增加,血浆 TGFβ 水平下降。此外,在追踪至治疗结束的14名参与者中(治疗开始后44至315天不等),有9名参与者表现出ISG、抗病毒限制因子和Toll样受体(TLR)信号通路的早期诱导和持续表达,以及HIV病毒库的减少。将这些转录组特征映射到1000多个公共多组学数据集上,结果显示抗PD-1诱导的程序存在于不同疾病状态的亚群中,表明部分人群已经表现出增强的抗病毒状态。总之,这些发现定义了有助于识别最有可能通过抗PD-1疗法实现病毒库衰减的HIV感染者的免疫通路,并提示持续的ISG激活可能有助于减少病毒库,并防止抗逆转录病毒疗法中断后病毒反弹。
+ + + + +



