

 English
English文献解读|Nat Genet(29):肿瘤免疫微环境的多模态空间表征可识别弥漫性大B细胞淋巴瘤中可靶向的炎症微环境
✦ +
+
论文ID
原名:Multi-modal spatial characterization of tumor immune microenvironments identifies targetable inflammatory niches in diffuse large B cell lymphoma
译名:肿瘤免疫微环境的多模态空间表征可识别弥漫性大B细胞淋巴瘤中可靶向的炎症微环境
期刊:Nature Genetics
影响因子:29
发表时间:2025.10.21
DOI号:10.1038/s41588-025-02353-5
背 景
弥漫性大B细胞淋巴瘤(DLBCL)是一组常见的、异质性的成熟B细胞恶性肿瘤,通常起源于淋巴结。淋巴结中基质细胞、抗原呈递细胞和其他髓系细胞共同促进T细胞和B细胞对病原体的适应性免疫应答。淋巴结的结构排列有利于复杂的细胞间相互作用,从而控制细胞迁移、增殖、克隆选择和负反馈,而这些对于强大的免疫应答及其感染后的清除至关重要。在DLBCL中,这种结构遭到破坏,恶性B细胞侵蚀了淋巴结的大部分区域,许多底层细胞类型仍以不同频率存在,并参与疾病的发生发展。虽然大多数DLBCL对一线化疗免疫疗法有反应,但仍有约40%的患者出现耐药或复发。这些患者的预后以往较差,但随着CD19嵌合抗原受体(CAR)T细胞以及近期双特异性抗体的引入,预后已显著改善。这些免疫疗法在部分患者中取得突破性成功,但在另一些患者中疗效有限,凸显了理解淋巴瘤免疫生物学以寻找改善患者预后策略的重要性。
实验设计
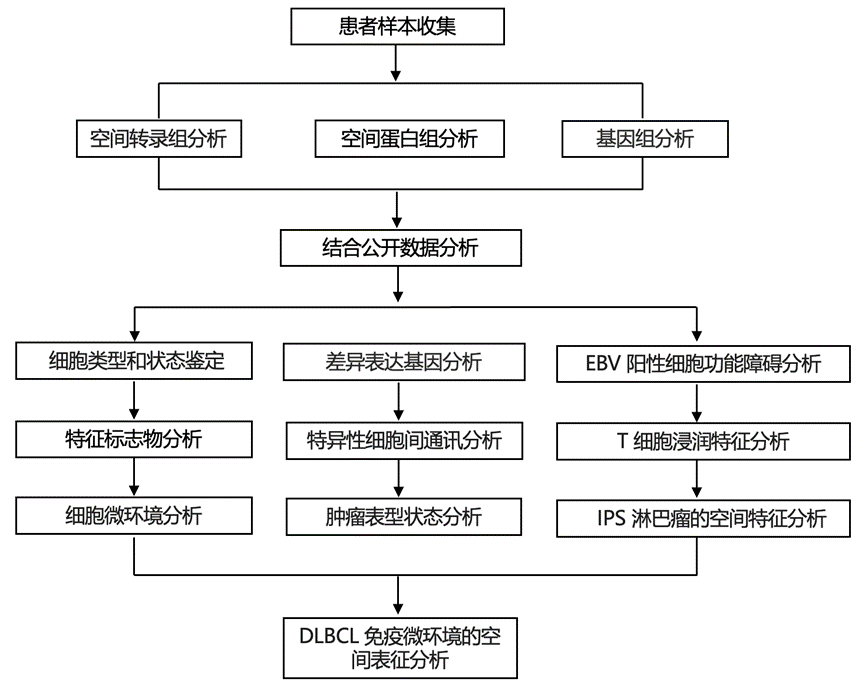
结 果
01

通过单细胞空间转录组学进行高分辨率细胞分型
研究团队从78例大B细胞淋巴瘤和5例对照(4例扁桃体,1例淋巴结)的切除活检组织中构建了6个组织微阵列(TMA)。B细胞淋巴瘤包括66例未另行分类的DLBCL;5例未另行分类的EB病毒阳性DLBCL;4例富含T细胞/组织细胞的大B细胞淋巴瘤;2例原发性纵隔大B细胞淋巴瘤;以及1例移植后淋巴增殖性疾病。其中,47例为初治病例,31例为复发/难治性病例。对75个样本的进行全外显子组测序分析,每个TMA均进行单细胞空间转录组学分析,并使用31-抗体共检测 InDEXing (CODEX) panel 进行空间蛋白质组学分析(图1a)。
经过严格的质量控制,从 CosMx 分析中保留了1322740个高质量、可表征的细胞用于后续分析。无监督聚类分析根据经典标记基因(图1c)鉴定出七种主要细胞类型(图1b)。每个TMA中的细胞分布于所有细胞类型中。为了验证细胞分型的准确性,他们使用了内部构建的大 B 细胞淋巴瘤单细胞核多组学 (snMultiome) 数据集,该数据集包含所有主要谱系。本研究的 CosMx 数据中的主要谱系(B 细胞、T 细胞、髓系和非造血细胞)在结合 snMultiome 数据的特征表达方面表现出高度特异性,并且在两个数据集的联合嵌入中具有共比对性。他们进一步使用蛋白质组学方法,在相同的TMA上(而非连续切片上)通过比较 CODEX 核心区域所有细胞和 CosMx 视野 (FOV) 中所有细胞的细胞频率,验证了细胞分型性能(图1d)。虽然CosMx 和 CODEX 在Z轴方向上分离,这阻碍了细胞水平的映射,但使用 CODEX 基于标记的方法识别出的四个主要谱系与 CosMx 鉴定出的细胞类型显示出显著相关性。在CODEX数据库中,B细胞和T细胞谱系的相关性最强,这由三个抗体标记物定义。两种方法均证实了不同样本间细胞组成和组织结构的高度异质性(图1d)。因此,利用CosMx平台进行空间转录组分析是一种强大的工具,可用于探索单细胞分辨率下的空间组成和基因表达。
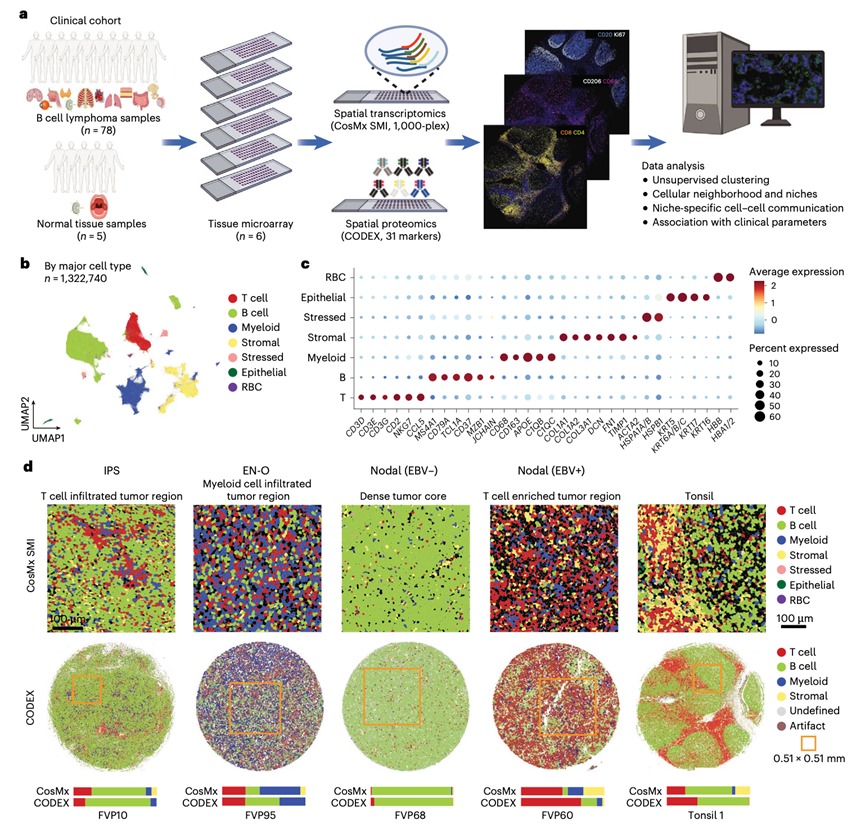
图1. CosMx SMI 和 CODEX 中的研究设计和主要细胞类型鉴定。
(a) 工作流程示意图。(b) CosMx SMI 数据集中识别的主要细胞类型的均匀流形近似和投影 (UMAP) 图。(c) 每种细胞类型的典型标记基因。(d) 来自五种不同临床状况的样本的代表性 CosMx SMI(和 CODEX图像。
02
DLBCL 包含空间上不同的 CN
对 CosMx 数据进行高分辨率聚类分析,确定了 19 种细胞类型和状态(图2a),其中包括一个占主导地位(35.27% 的细胞)的 B 细胞聚类,其特征是增殖标志物(TOP2A、PCNA)高表达以及免疫球蛋白重链和轻链表达受限,因此可能代表肿瘤 B 细胞(C0_Tumor-B)。他们还鉴定了三个浆细胞聚类,它们以不同的免疫球蛋白重链表达为特征(C1_PC_IgG、C2_PC_IgA 和 C4_PC_IgM),以及一个表达 CD44 的静息 B 细胞群(C3_Resting-B)。定义了一个单一的T细胞聚类(C5_T),以及肿瘤相关巨噬细胞(TAM)(C6_TAM_APOE_C1Q 、 C7_TAM_SPP1)和其他多种巨噬细胞亚群(C8_Mac_DUSP1、C9_Mac_CXCL8、C10_Mac_MT2A )。非造血成分包括多种亚群,例如滤泡网状细胞(FRC)(C11_FRC)、高内皮静脉细胞(C12_HEV)、表达血管性血友病因子(VWF)的内皮细胞(C13_Endothelial_VWF)、血管平滑肌细胞(C14_VSMC)、滤泡树突状细胞(C15_FDC)和少量上皮细胞(C17_Epithelial)。最后,他们检测到红细胞(C18_RBC)和表达热休克蛋白的细胞(C16_Stressed)聚类,后者是细胞应激的标志。
单细胞空间转录组学相对于单细胞转录组学分析(scRNA-seq)的一个关键优势在于能够将细胞映射回二维空间,从而识别细胞邻居和细胞微环境(CN)。事实上,过去十年空间研究积累的知识,使人们越来越认识到组织特化细胞群是作为一个功能单元协同工作的,而非各自独立运作。基于此,他们对每个细胞进行了细胞邻域分析,并随后利用空间统计方法分析了免疫细胞彼此之间以及与肿瘤细胞之间建立功能性联系的能力。细胞邻域定义为质心位于邻域中心200像素范围内的细胞集合(图2b)。不同半径下的邻域结构高度相似。因此,他们选择了200像素的阈值,该阈值下每个邻域包含约10-20个细胞,以与之前的研究保持一致。因此,邻域组成分析表明,以相同细胞状态为中心的邻域中,该细胞状态总是高度富集(图2c)。相似细胞的空间聚集倾向提示了不同细胞邻域的形成。因此,通过对细胞邻域进行聚类分析,将空间组织模式可重复地表征为微环境(图2d)。这确定了七种典型的 CN,其特征为 T 细胞富集(CN1_T;占细胞总面积的 8.76%)、浆细胞富集(CN2_PC;占肿瘤总面积的 2.34%)、髓系细胞富集(CN3_Myeloid;占肿瘤总面积的 9.18%)、基质细胞富集(CN4_Stromal;占肿瘤总面积的 6.93%)、肿瘤 B 细胞富集(CN5_Tumor-B;占肿瘤总面积的 14.21%)、肿瘤细胞弥漫分布于异质性免疫浸润中(CN6_Diffuse;占肿瘤总面积的 35.47%)或非恶性细胞与散在肿瘤细胞混合分布(CN7_Mixed;占肿瘤总面积的 23.12%)(图2e)。CN1-CN5 中各细胞状态的优势体现在其细胞状态特异性差异表达基因 (DEG) 中,而 CN6 和 CN7 的DEG则呈现混合模式,反映了其多样化的细胞组成(图2f)。他们展示了每个细胞微环境及其对应细胞状态的代表性视野 (FOV)(图2g)。此外,使用来自相同样本的正交 CODEX 图像验证了 CN1-CN5 中主要细胞类型(T 细胞或自然杀伤细胞、B 细胞、髓系细胞和非造血细胞)的空间聚集情况。在每个特定的细胞微环境(CN1-CN5)中,优势细胞状态占据了细胞共定位网络的中心,但在混合细胞微环境(CN7)中未观察到明显的中心。有趣的是,在弥散细胞微环境(CN6)中,肿瘤B细胞形成了共定位网络的中心,表明该微环境中肿瘤B细胞与多种免疫细胞类型之间存在接触。肿瘤B细胞比例最高的两个微环境是CN5和CN6,分别代表肿瘤细胞密集分布和肿瘤细胞弥散分布于免疫浸润细胞中(图2e)。在78个肿瘤中,有75个肿瘤存在不同程度的 CN6 弥散细胞微环境,中位数为 33% 的细胞;而 CN5 致密肿瘤-B 细胞微环境则更为局限,在 78 个肿瘤中仅有 58 个肿瘤观察到,中位数为 5% 的细胞。
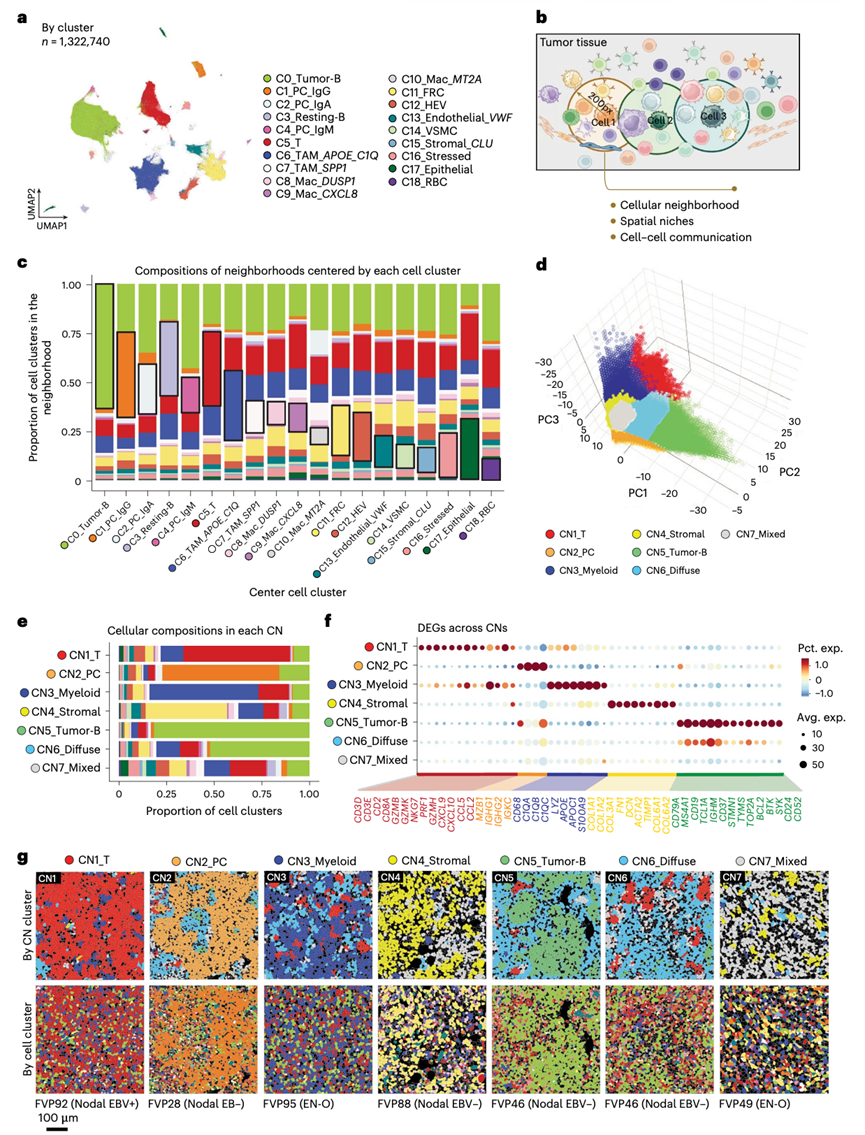
图2. B 细胞淋巴瘤中的细胞邻域结构和独特的空间生态位。
(a) CosMx SMI 数据集中的高分辨率细胞状态识别。(b) 细胞邻域分析示意图。(c) CosMx SMI 数据集中 19 种不同细胞状态的邻域组成。(d) 基于细胞邻域组成的细胞无监督聚类,鉴定出七个独特的空间生态位。(e) 每个空间生态位的细胞状态组成。(f) 每个空间生态位中的代表性差异表达基因 (DEG) 。(g) 来自 CosMx SMI 的代表性视野 (FOV) 图像,每个图像均具有一个主要的空间生态位。
03
微环境特异性细胞间通讯
虽然初始阶段的无监督聚类仅定义了一个T细胞聚类(C5_T),但他们推测,由于相邻细胞间通讯模式的差异,T细胞表型在不同的细胞微环境结构中可能存在差异。为了验证这一可能性,他们对CN1(富含T细胞)、CN3(富含髓系细胞)和CN5(富含肿瘤B细胞)中的T细胞基因表达差异进行了有监督分析。结果显示,CN1中的T细胞表达最高水平的幼稚/记忆T细胞标志物、高水平的细胞毒性T细胞标志物和低水平的功能障碍T细胞标志物(图3a-b),表明其T细胞功能更完善、更健康。相比之下,CN3 内的T细胞幼稚/记忆标志物的表达较低,但细胞毒性标志物的表达水平升高(图3a),同时LAG3、HAVCR2 (TIM3) 和TNFRSF9 (4-1BB) 的表达也较高,表明其具有活化的效应表型,并伴有功能障碍或耗竭的迹象。CN5 内的 T 细胞细胞毒性标志物的表达并不强烈,而是表达显著更高水平的其他功能障碍标志物,例如ENTPD1、TIGIT和PDCD1,提示肿瘤富集的微环境对 T 细胞功能具有抑制作用(图3a)。为了研究 T 细胞募集和排斥的机制,他们采用了两种方法。首先,评估了各微环境DEG中T细胞趋化因子的表达情况,结果显示,在富含T细胞的CN1和富含髓系细胞的CN3中,多种细胞类型均高表达多种趋化因子,包括APOE +C1Q+肿瘤相关巨噬细胞(TAM)(C6)、肿瘤B细胞(C0)和滤泡网状细胞(FRC)(C11),但在富含肿瘤细胞的CN5中未观察到(图3b-c)。此外,富含T细胞的微环境(CN1)中的APOE+C1Q+ TAM(C6)也表达最高水平的抗原呈递基因(图3d)。这可能有助于改善该微环境中观察到的T细胞健康状况。第二种方法是细胞间通讯分析,该分析通过证实T细胞富集微环境(CN1)内多种趋化因子-趋化因子受体相互作用的显著富集,支持了先前的结果。这些趋化因子来源于多种细胞类型,相应的趋化因子受体表达于T细胞上。在其他细胞微环境中,包括肿瘤富集微环境(图3e),均未观察到此类相互作用。对相邻的APOE +C1Q+ TAM和T细胞(图3f)或高内皮静脉细胞和T细胞内单分子分辨率转录表达的分析清晰地显示了相邻细胞上的趋化因子和趋化因子受体表达,支持了每个微环境内存在局部相互作用的观点。细胞间通讯分析也证实,在富含T细胞的微环境中,MHC II类分子(MHC-II)介导的抗原呈递增强,CD4 T细胞的抗原呈递增加。相反,在富含髓系细胞和富含肿瘤细胞的微环境中,CD274 - PDCD1(PD-L1-PD-1)相互作用增强,PD-L1分别来源于髓系细胞和肿瘤细胞(图3g-h)。综上所述,这些发现突显了不同CN中T细胞状态和表型的显著差异,这可能受到各微环境中不同的细胞间通讯模式的影响。
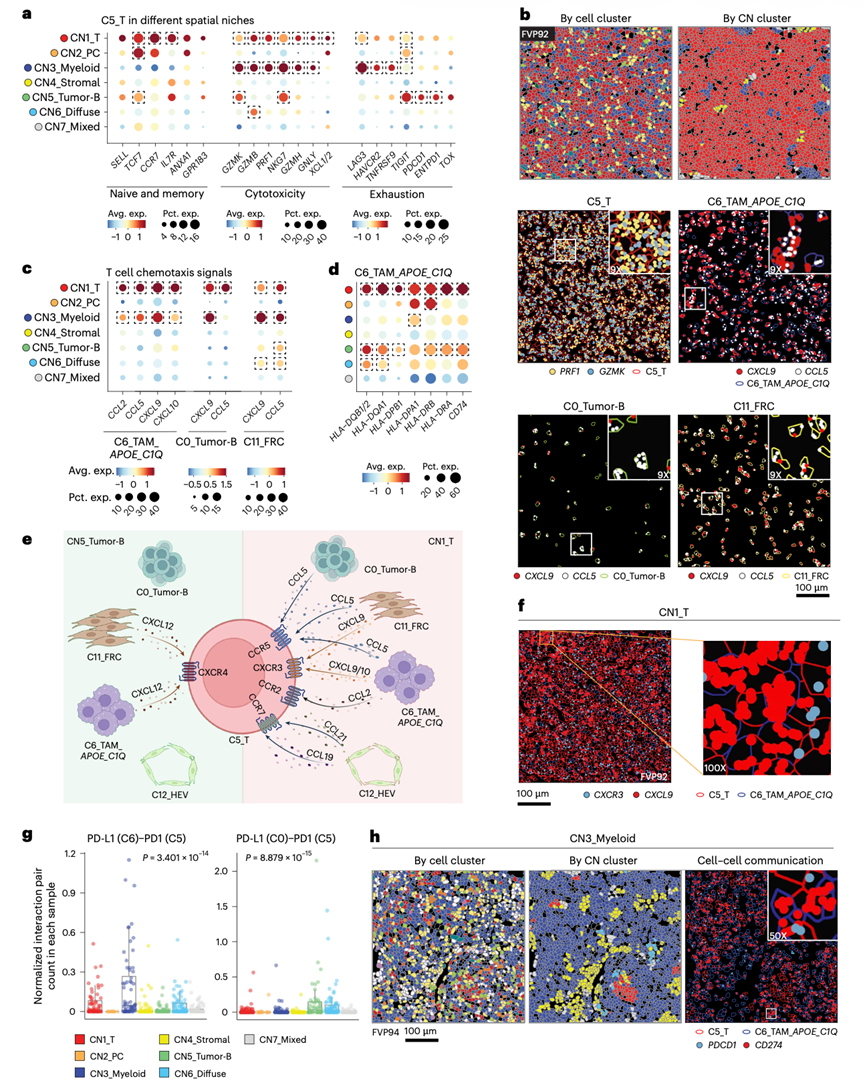
图3. T 细胞的微环境特异性功能状态。
(a) 不同空间微环境中T细胞中幼稚/记忆、细胞毒性和耗竭标志物的表达。(b) CosMx SMI中以CN1为主的代表性视野。(c) 来自不同空间微环境中的APOE+C1Q+ TAM、肿瘤B细胞和FRC中的T细胞趋化分子。(d) MHC-II基因的表达。(e) CN1和CN5中T细胞趋化通路的总结。(f) 胞中CXCL9和CXCR3的表达。(g) 柱状图描绘了每个空间微环境中归一化的 PD-L1-PD-1 相互作用计数。(h) TAM 和 T 细胞之间的 PD-L1-PD-1 相互作用。
04
肿瘤B细胞微环境反映了生发中心相互作用
在所有CN中均发现了肿瘤B细胞,但肿瘤B细胞在CN5中最为富集,占据了大部分空间区域;而在CN6中,肿瘤B细胞则与多种免疫浸润细胞混合存在。为了探究导致这些空间微环境差异的机制,他们检测了CN5和CN6中肿瘤B细胞的特征。他们观察到,与CN6中的肿瘤B细胞相比,CN5中的肿瘤B细胞表达更高水平的细胞周期相关标志物和抗凋亡癌基因BCL2 (图4a-b),这进一步证实了肿瘤B细胞的快速扩增可能导致了这些区域中观察到的空间消失现象。对DEG的监督分析发现,与CN6驻留肿瘤B细胞相比,CN5驻留肿瘤B细胞中多个与淋巴瘤生物学相关的基因表达显著升高,包括参与BCR信号通路和表观遗传调控的基因。细胞间通讯分析揭示了CN5和CN6微环境之间的其他差异。在肿瘤富集的CN5微环境中,CXCL12-CXCR4相互作用最为显著,其中FRC为表达CXCR4的肿瘤B细胞提供CXCL12(图4c-d),而FRC和APOE+C1Q+TAM则共同为T细胞提供CXCL12(图4e-f)。相比之下,CN1 内的肿瘤 B 细胞与 CD4 T 细胞表现出显著的 CD40L-CD40 相互作用,其中一部分 B 细胞还高表达多种免疫检查点配体。因此,主要的肿瘤 B 细胞微环境呈现出类似于生发中心暗区的模式。暗区受 CXCR4 调控,聚集着高度增殖的 B 细胞,与生发中心亮区相比,其 T 细胞数量较少。在生发中心亮区,B 细胞增殖能力较低,并接受来自 CD4 T 细胞的 CD40 介导的共刺激信号。
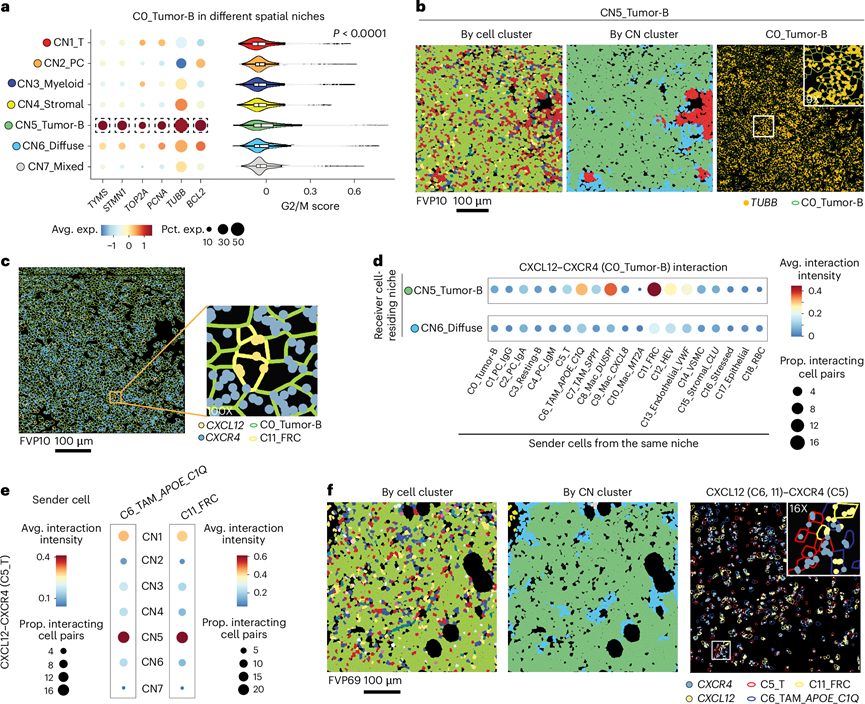
图4. CN5 中独特的肿瘤表型状态和细胞相互作用。
(a) 不同空间微环境中肿瘤B细胞的细胞周期标志物表达及G2/M期特征评分。(b) 代表性视野(FOV)显示CN5肿瘤B细胞中细胞周期标志物TUBB的广泛表达。(c) 与图(b)相同视野中FRC与肿瘤B细胞的CXCL12-CXCR4相互作用。(d) CN5和CN6中所有肿瘤微环境(TME)细胞与肿瘤B细胞的CXCL12-CXCR4相互作用活性。(e) TAM或FRC与T细胞的CXCL12-CXCR4相互作用活性。
05
EBV阳性弥漫性大B细胞淋巴瘤中的T细胞功能障碍
EBV是一种B细胞转化病毒,约5%的DLBCL病例是由其引起的。在健康个体中,EBV感染的B细胞受到T细胞的监视,从而抑制恶性增殖。因此,他们利用临床EBER原位杂交检测获得的EBV状态,比较了免疫功能正常个体(即排除移植后淋巴增殖性疾病;n = 5)的淋巴结EBV阳性DLBCL与淋巴结EBV阴性DLBCL的T细胞特征。结果显示,EBV阳性DLBCL中肿瘤B细胞(C0)的比例显著降低,而T细胞(C5)的比例显著升高(图5a)。因此,在EBV阳性DLBCL中,肿瘤富集的CN5微环境仅占细胞总数的0.04%,而T细胞富集的CN1微环境和混合的CN7微环境则显著过度表达(图5b),且T细胞主要存在于这些微环境中(图5c-d)。与此观察结果一致,细胞间通讯分析显示,在EBV阳性DLBCL中,T细胞趋化信号更强,且通过MHC-I和MHC-II接收抗原信号。这包括与肿瘤B细胞更密切的相互作用,在EBV阳性病例中,肿瘤B细胞更频繁地与T细胞和APOE+C1Q+ TAM相邻,而EBV阴性病例中则较少与其他肿瘤B细胞相邻( 图5e),正交CODEX数据也证实了这一点。鉴于EBV阴性和EBV阳性DLBCL中CN的比例存在差异,并且由于微环境驻留与功能差异相关,他们通过将EBV阴性和EBV阳性肿瘤之间T细胞功能的比较限制在T细胞微环境(CN1)内,来控制这一混杂因素。尽管在EBV阴性DLBCL中,T细胞微环境(CN1)支持初始或记忆T细胞,且这些T细胞的耗竭标志物表达水平较低,但在EBV阳性DLBCL中,细胞毒性和耗竭标志物的表达水平显著升高(图5f-i)。通过CosMx在单RNA分子分辨率下(图5j)以及通过CODEX蛋白染色(图5k)均证实了这一点。可以清楚地观察到,EBV阳性DLBCL中CN1 T细胞微环境中的T细胞表现出显著更高的效应分子表达。此外,EBV阳性DLBCL中的T细胞共表达HAVCR2等耗竭标志物(图5j)。综上所述,这些结果表明,虽然EBV阳性DLBCL中T细胞和混合微环境的过度表达(这些微环境通常有利于EBV阴性DLBCL中的T细胞功能),但EBV阳性肿瘤中的T细胞却表现出细胞毒性和耗竭标志物,表明其处于慢性刺激和功能障碍状态。
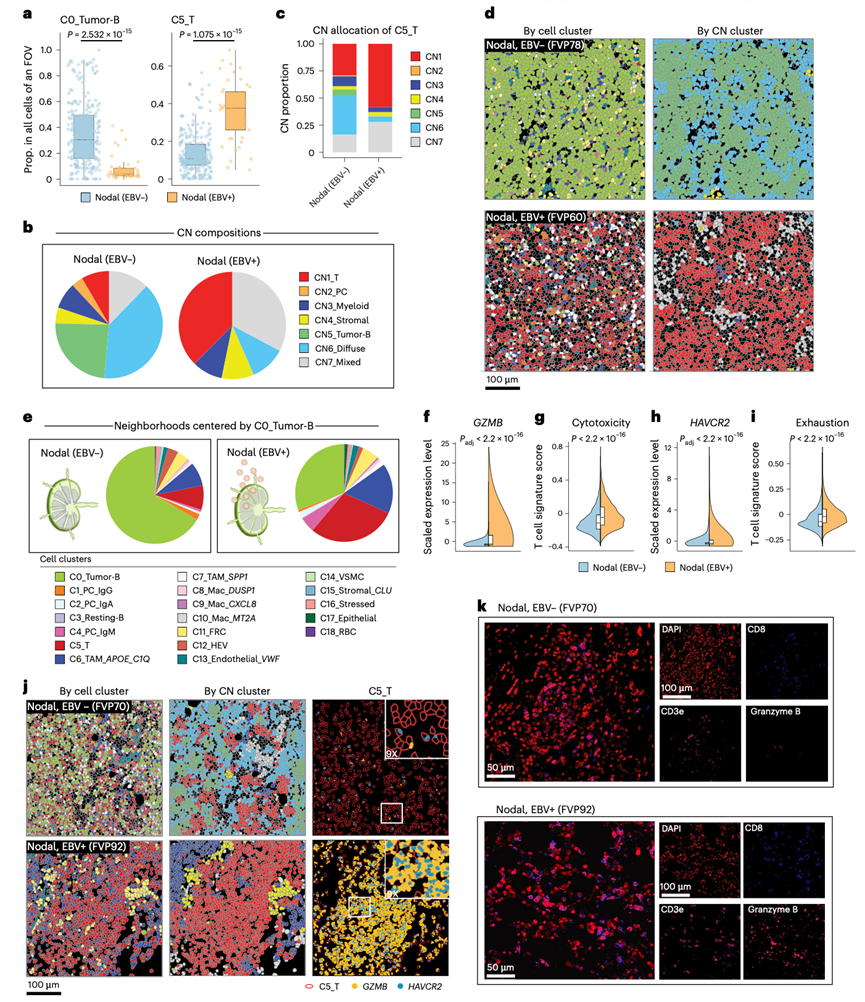
图5. 淋巴结 B 细胞淋巴瘤的空间分辨细胞图谱。
(a) 比较EBV阳性和EBV阴性淋巴结B细胞淋巴瘤中肿瘤B细胞和T细胞的细胞比例。(b) 饼图表示EBV阳性和EBV阴性淋巴结病变的CN组成。(c) 分段条形图描绘了不同EBV感染状态的淋巴结病变中T细胞的CN分布。(d) CosMx SMI的代表性视野(FOV)显示了EBV阳性和EBV阴性淋巴结病变中不同的细胞组成。(e) EBV阳性和EBV阴性淋巴结病变中肿瘤B细胞的邻域结构。(f-i) EBV阳性和EBV阴性淋巴结病变中T细胞代表性细胞毒性或耗竭标志物和特征的表达。(j) CosMx SMI的代表性FOV图像显示了EBV阳性和EBV阴性淋巴结病变中T细胞GZMB和HAVCR2的表达。(k) 代表性 CODEX 图像。
06
免疫豁免部位弥漫性大B细胞淋巴瘤中的T细胞浸润
虽然DLBCL起源于中枢神经系统(CNS)和睾丸等免疫细胞迁移受限的部位,但免疫豁免部位(IPS)的DLBCL也发现存在免疫细胞浸润。通过比较11例IPS DLBCL(CNS,n = 5;睾丸,n = 5;眼部,n = 1)与28例淋巴结DLBCL和23例其他结外部位(EN-O)DLBCL,探讨了数据集中的IPS DLBCL的免疫学特征。脾脏和EBV阳性病例排除在本分析之外。虽然IPS DLBCL中肿瘤B细胞(C0)的比例高于淋巴结和EN-O DLBCL,但T细胞(C5)的频率无显著差异(图6a)。APOE +C1Q +TAM (C6) 和 FRC (C11) 在 EN-O DLBCL 中更为丰富,但在淋巴结 DLBCL 和 IPS DLBCL 之间无显著差异(图6a)。IPS 和 EN-O DLBCL 中,肿瘤富集的 CN5 微环境中的细胞比例均较低,而弥散的 CN6 和混合的 CN7 微环境中的细胞比例较高(图6b)。淋巴结 DLBCL 中,肿瘤富集的 CN5 微环境中相邻的肿瘤 B 细胞比例较高(图6c)。虽然位于免疫豁免部位,IPS DLBCL 中的肿瘤 B 细胞与 T 细胞的相邻程度与淋巴结 DLBCL 大致相同,且显著高于 EN-O DLBCL(图6d)。此外,淋巴结和IPS DLBCL中,大致相同比例的T细胞位于弥漫性CN6微环境中,其中T细胞和其他细胞与肿瘤B细胞混合存在;而在EN-O DLBCL中,更多比例的T细胞位于混合的CN7微环境中,其中肿瘤B细胞较少(图6e-f)。此外,他们观察到细胞毒性特征表达显著升高(图7a),表达效应分子(GZMB和PRF1)(图7b)的细胞比例更高,以及增殖率更高(图7c-g)。与淋巴结或EN-O DLBCL相比,IPS DLBCL中T细胞的表达水平显著升高,提示IPS DLBCL内存在持续的抗原反应和T细胞扩增。然而,IPS DLBCL中T细胞也表现出与T细胞功能障碍相关的标志物(如LAG3、HAVCR2、PDCD1和ENTPD1)的高表达(图7h-i)。此外, IPS DLBCL中的其他细胞类型也高表达CD274 (PD-L1)等配体(图7j),并且IPS DLBCL中存在显著的Galectin-9-TIM3(HAVCR2)信号通路(图7k)。这些共同表明IPS DLBCL中存在免疫抑制性微环境。这些数据共同表明,IPS DLBCL 在弥漫和混合 CN 中具有强大的 T 细胞浸润,具有强烈的增殖和细胞毒性,但也伴有共抑制信号,这突显了 IPS DLBCL 可能适合免疫疗法。
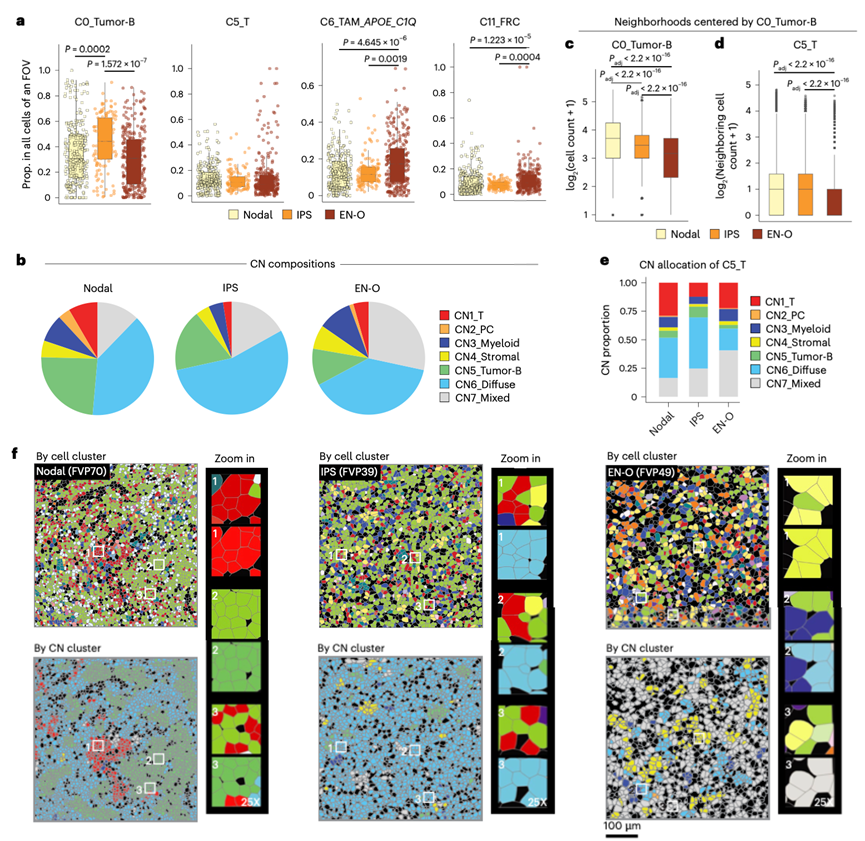
图6. 在不同解剖部位发生的 B 细胞淋巴瘤的空间分辨细胞景观。
(a) 不同解剖部位(包括淋巴结、间质肉瘤和结外病变)发生的B细胞淋巴瘤的细胞组成。(b) 不同解剖部位肿瘤的拷贝数组成饼图。(c-d) 不同解剖部位肿瘤B细胞及其周围T细胞群的箱线图。(e) 不同肿瘤部位T细胞拷贝数分布的子条形图。(f) CosMx SMI 的代表性视野显示了不同解剖部位肿瘤中不同的细胞组成。
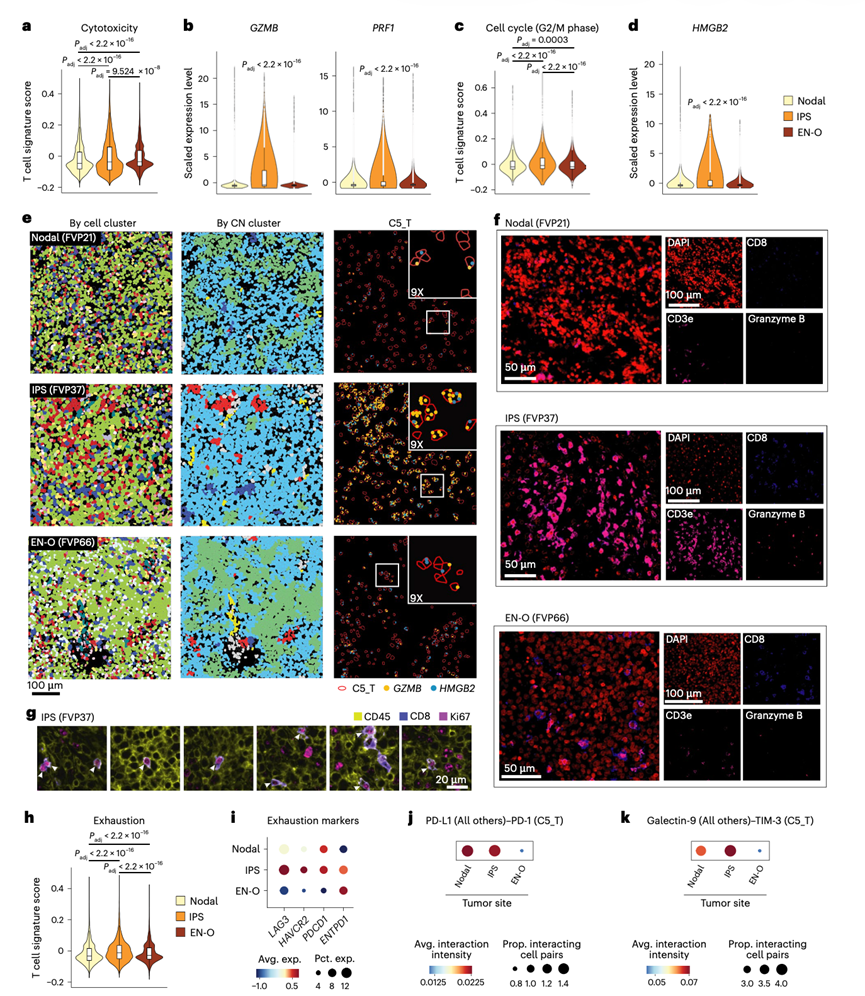
图7. IPS 淋巴瘤的空间特征。
(a) 不同肿瘤部位淋巴瘤中细胞毒性标志物的表达。(b) 淋巴结、IPS和EN-O病变中T细胞中细胞毒性标志物GZMB和PRF1的表达。(c) 根据肿瘤部位分析G2/M细胞周期标志物的表达。(d) 根据肿瘤部位分析代表性细胞周期基因HMGB2的表达。(e) CosMx SMI 的代表性视野显示了细胞毒性标志物GZMB和增殖标志物HMGB2在不同肿瘤部位 T 细胞中的表达。(f-g) 与图(e)相同样本的代表性 CODEX 图像。(h-i) 不同肿瘤部位 T 细胞中耗竭特征和代表性耗竭标志物的表达。(j-k) 不同肿瘤部位所有其他细胞与 T 细胞之间的 PD-L1-PD-1 相互作用和 Galectin-9-TIM3 相互作用。
+ + + + + + + + + + +
结 论
本研究采用高通量空间转录组学和蛋白质组学以及基因组分析,对78例DLBCL肿瘤的免疫微环境结构进行了表征,定义了七个不同的CN,每个微环境都具有独特的细胞组成、空间组织和细胞间通讯模式,并伴有微环境特异性的T细胞和肿瘤B细胞表型。其中,来自免疫豁免部位的DLBCL在弥漫性微环境中显示出大量的T细胞浸润,免疫细胞与肿瘤B细胞混合存在,并具有激活和效应功能的转录特征,提示这些微环境可能已启动抗肿瘤免疫反应。因此,对 DLBCL 免疫微环境的空间表征揭示了CN,这些CN促进了不同的细胞间相互作用模式,从而导致了微环境内肿瘤细胞和免疫细胞的表型异质性。
+ + + + +



