

 English
English文献解读|Nat Commun(15.7):早期人类嗅觉发育的单细胞和空间图谱
✦ +
+
论文ID
原名:A single-cell and spatial atlas of early human olfactory development
译名:早期人类嗅觉发育的单细胞和空间图谱
期刊:Nature Communications
影响因子:15.7
发表时间:2026.04.17
DOI号:10.1038/s41467-026-71595-6
背 景
嗅觉由人类嗅觉系统控制,是人类感知外部环境的重要组成部分,有助于人类与复杂环境互动。它在进食、识别有害气味和社交关系等重要功能中发挥着关键作用。嗅觉丧失会严重影响人的生活质量,并可能导致食欲下降、营养不良,甚至焦虑或抑郁等神经系统紊乱。嗅觉感觉神经元(OSN)是主要嗅觉上皮内的主要检测器,在强大的选择压力下进化,能够识别超过10000种不同分子的气味化学空间。气味分子由一类与G蛋白偶联受体相关的跨膜受体检测,这类受体占功能基因总数的50%以上,小鼠有超过1200个嗅觉受体(OR)基因,人类约有400个。每个OSN以单基因和单等位基因的方式表达单个OR基因。对特定OR的表达受多种机制调控。人类鼻腔区域起源于神经嵴和鼻基板谱系,但由于胎儿组织获取有限且结构复杂,其早期发育过程仍知之甚少。
实验设计
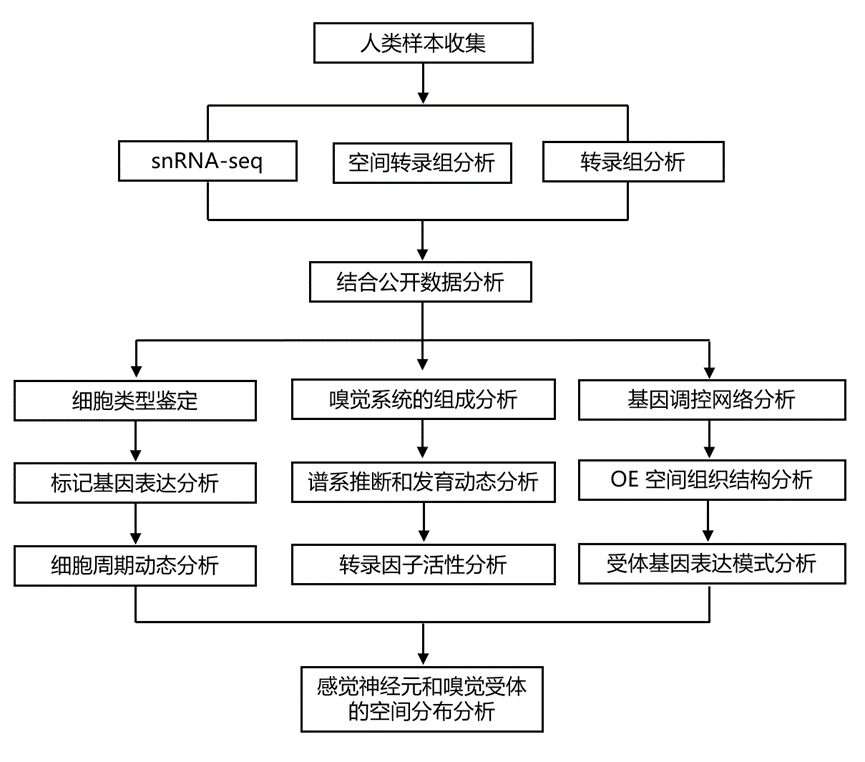
结 果
01

人类胎儿鼻腔单核图谱
为了表征发育中人类鼻腔区域的细胞图谱,研究团队对8个胎儿样本(孕周7-12周)(图 1a)进行了单细胞核转录组分析(snRNA-seq)。
在对样本进行解复用并对单个细胞核进行供体分配后(图1b),整合了所有发育阶段的数据集,生成了包含41875个细胞核的统一转录图谱(图1c)。大多数细胞核分类为单细胞核,少数鉴定为双细胞核或未分配。经过质量控制过滤后,保留了一组平衡的、高质量的供体匹配的细胞核用于后续分析。性别判定采用每个细胞核内XIST与Y染色体连锁基因表达比值,将细胞核分为男性样、女性样或性别模糊。在质量控制过程中,剔除了一小部分性别特征不一致且红系基因表达较弱的样本,这与母体血液来源的微量RNA相符。基因表达相关性的层次聚类将胎儿鼻部区域分解为主要细胞区室,包括上皮细胞、神经元、间充质细胞、血管细胞、免疫细胞、肌肉细胞和神经嵴衍生的胶质细胞群。
为了评估细胞组成的动态变化,他们定量了不同发育阶段的细胞类型比例。间充质细胞群表现出显著的动态变化,多种间充质和成骨亚型随时间推移而增加,这与活跃的颅面生长和组织重塑相一致;而神经嵴细胞(NCC)、GnRH神经元和NOS1神经元则呈现相反的趋势。促性腺激素释放激素(GnRH)神经元调节哺乳动物的生育能力,它们起源于嗅觉基板,并在胎儿早期发育过程中沿着终末神经和犁鼻神经迁移至大脑。在妊娠第6-7周(PCW6-7),这些神经元与源自基板和神经嵴的迁移团(MM)细胞一起迁移穿过鼻区。与此过程一致,他们观察到在妊娠第7周(PCW7)至第12周(PCW12)之间,鼻区GnRH神经元和NOS1+ MM成分显著减少,这反映了它们逐渐迁移至大脑的过程。两种不同注释级别的UMAP嵌入图显示了不同细胞实体的一致组织结构。由此产生的无监督聚类和注释结果识别出32种不同的细胞群(图1d),这与先前在其他模型中的研究结果一致。神经元和胶质细胞群表现出高度的转录多样性,包括嗅觉神经元、神经祖细胞、神经嵴细胞及其衍生物,以及雪旺氏细胞和嗅鞘细胞(OEC)(图 1d-e)。非神经元细胞群包括呼吸道上皮细胞、间充质基质细胞、周细胞、软骨和成骨细胞谱系、内皮细胞、骨骼肌细胞和卫星细胞(图1d-e)。他们还鉴定出丰富的免疫细胞群,提示鼻腔区域与胎儿免疫发育之间可能存在联系。在整个发育阶段,未观察到细胞类型组成存在性别差异(图1e-f)。
接下来,他们对不同功能类别中的标记基因表达进行了可视化分析。他们观察到上皮细胞、神经嵴衍生物和神经元、间充质细胞和软骨细胞、血管细胞、肌肉细胞和免疫细胞谱系均具有独特的基因特征(图1g),从而能够对发育中的嗅觉系统中的细胞类型进行可靠的分类。许多高表达基因代表了典型的细胞类型标记基因;然而,超过一半的基因此前未曾报道(图 1g)。
他们利用经典的基因表达谱,在嗅上皮(OE)中评估了早期人类鼻发育过程中的细胞周期动态。UMAP 嵌入分析显示,细胞按细胞周期阶段清晰分离,包括 G1 期、S 期、G2/M 期和有丝分裂后状态。对发育过程中细胞数量的定量分析表明,在 PCW7 至 PCW12 之间,活跃增殖(G2/M)细胞数量逐渐减少,同时 G1 期细胞数量增加,这与细胞从增殖向分化的转变相一致。细胞类型解析分析显示,增殖性基底细胞、球状基底细胞、雪旺氏细胞前体和间充质细胞群构成了大部分S期和G2/M期细胞,而包括未成熟嗅觉感觉神经元(iOSN)、直接神经元前体(INP)、谷氨酸能神经元(GLUT)、GABA能神经元(GABA)、促性腺激素释放激素(GnRH)神经元和NOS1神经元在内的神经元群则主要处于有丝分裂后状态。这些模式得到了经典细胞周期标志物表达和阶段评分的证实。
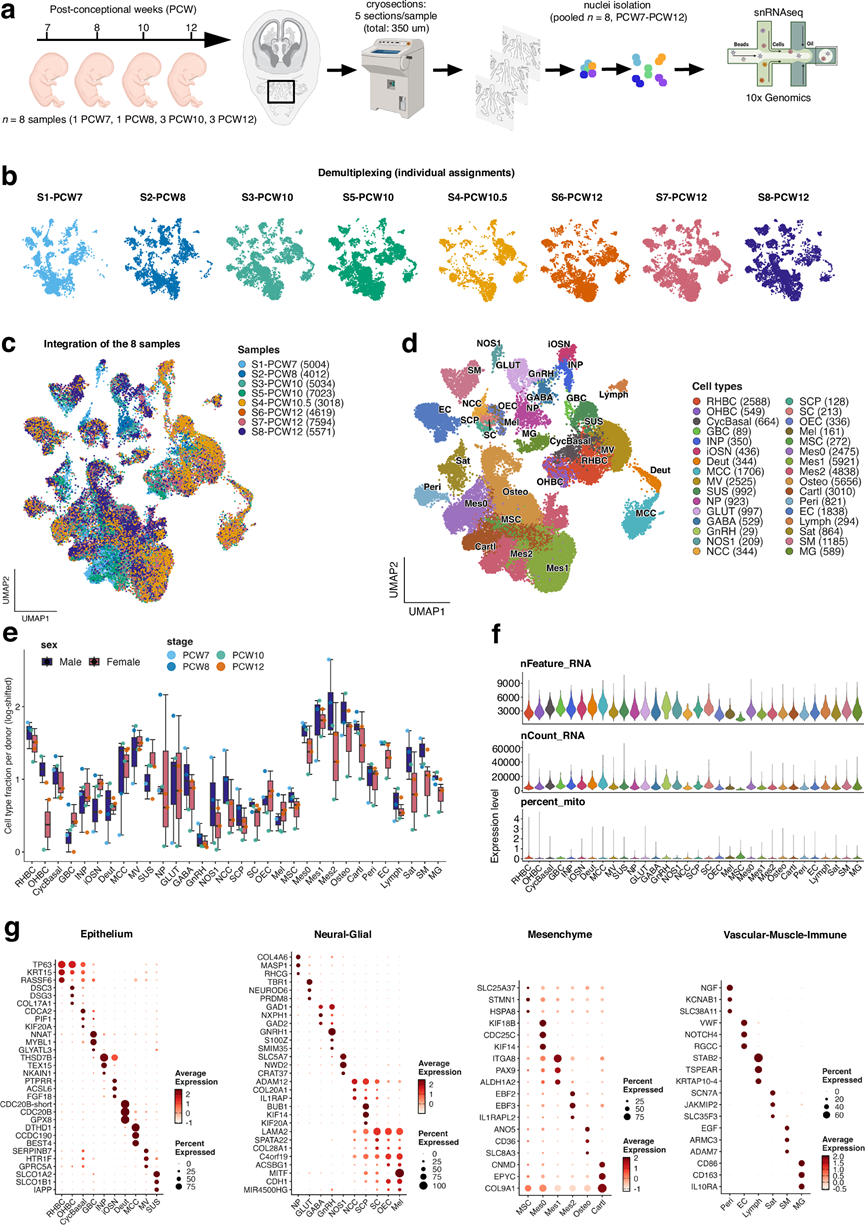
图1. 发育中的人类嗅觉感觉上皮的单细胞核转录组分析。
(a) 实验设计和snRNA-seq工作流程的示意图。(b) UMAP投影图,按样本来源着色。(c) 所有样本(PCW7-12)的整合UMAP嵌入图,按供体身份着色。(d) 整合数据集的UMAP投影图,按注释的细胞类型着色。(e) 按性别分层的样本水平细胞类型比例。(f) 小提琴图显示了注释细胞类型的质量控制指标,包括每个细胞核检测到的基因数(nFeature_RNA)、总UMI 计数(nCount_RNA)和线粒体转录本百分比。(g) 主要细胞类型中代表性标记基因表达的散点图,按功能类别分组。
02
人类胎儿嗅觉系统的组成和发育
接下来,他们将研究重点放在由嗅觉上皮和呼吸上皮组成的嗅觉系统上。他们结合已建立的小鼠和人类标记基因,确定了每个细胞聚类的细胞类型。他们鉴定了典型的嗅觉上皮细胞群,包括球状基底细胞(GBC)、水平基底细胞(OHBC)、中间核细胞(INP)、iOSN细胞、支持细胞(SUS)和微绒毛细胞(MV),以及相邻的呼吸上皮细胞群(图2a-b)。转录组分析揭示了基底细胞状态之间清晰的分子差异。GBC表达祖细胞相关基因,例如MYBL1、NNAT和HES6,而OHBC则富集MEG3、TP63和KRT5(图 2c-d)。值得注意的是,与 RHBC 相比,OHBC 中富集了MEG3,这是一种与上皮调节有关的长链非编码 RNA(图 2c-d),这与先前在人类成年 OE 中的发现一致。
对上皮细胞谱系丰度随发育时间变化的定量分析表明,CyclBasal、GBC 和 INP 显著减少,同时 MV 细胞和 iOSN 细胞数量增加,主要发生在 PCW7 至 PCW8 时期(图 2e)。OHBC 在最早的发育时间点(PCW7 和 PCW8)未出现,但在 PCW10 和 PCW12 时期出现(图 2e)。为了验证转录定义的嗅觉上皮细胞类型的空间组织和解剖学背景,他们在PCW9细胞中进行了免疫荧光和RNAscope实验(图 2f-g)。此外,基于包含294个基因的MERFISH标记基因,利用MERFISH进行空间转录组定位进一步验证了不同细胞群的解剖学定位(图S7a-b)。
研究表明,Olig2在小鼠的早期妊娠期至成年期,于OSN中瞬时表达,并在成熟的OSN中表达下调,从而促进OSN的最终成熟。与此一致,在妊娠第9周(PCW9),OLIG2免疫染色清晰地勾勒出OE的轮廓,并将其与相邻的呼吸上皮(RE)区分开来,证实了发育中鼻腔内的区域性分隔(图 2f)。他们还在呼吸上皮中检测到了MUC5AC的表达,以及GNRH1阳性神经元在鼻腔间充质中迁移(图 2g)。PCW7.5 时OE的透射电子显微镜显示其上皮超微结构,显示 OE 中存在 MV(粉红色)、SUS(绿色)、OSN(蓝色)和基底细胞(黄色),从而为早期嗅觉上皮特化提供了超微结构证据(图 2h)。
SOX2是神经干细胞和祖细胞的标志物,据报道在小鼠妊娠晚期和成年期,其在OE和RE的嗅觉干细胞以及顶端OE的支持细胞中表达。神经元和祖细胞标志物的免疫染色显示OE呈层状结构,基底层上方的TUJ1+神经元将iOSN轴突延伸至嗅球,而SOX2+ 祖细胞则定位于基底层和中间层以及整个RE(图 2i)。与转录组数据(图 2e)一致,在妊娠7.5周(PCW7.5)时,OE中未检测到OHBC,这由KRT5表达的缺失所证实,KRT5在该阶段仅限于RE表达(图 2i)。PCW12 时,OE 和 RE 中均存在 KRT5+ OHBC,且上皮厚度和结构存在显著差异(图2j)。SOX2 和 KRT5 的共染色证实了基底细胞在两种上皮中的基底膜定位,并显示 OE 中的 OHBC 呈层状、反应样形态排列(图 2j)。
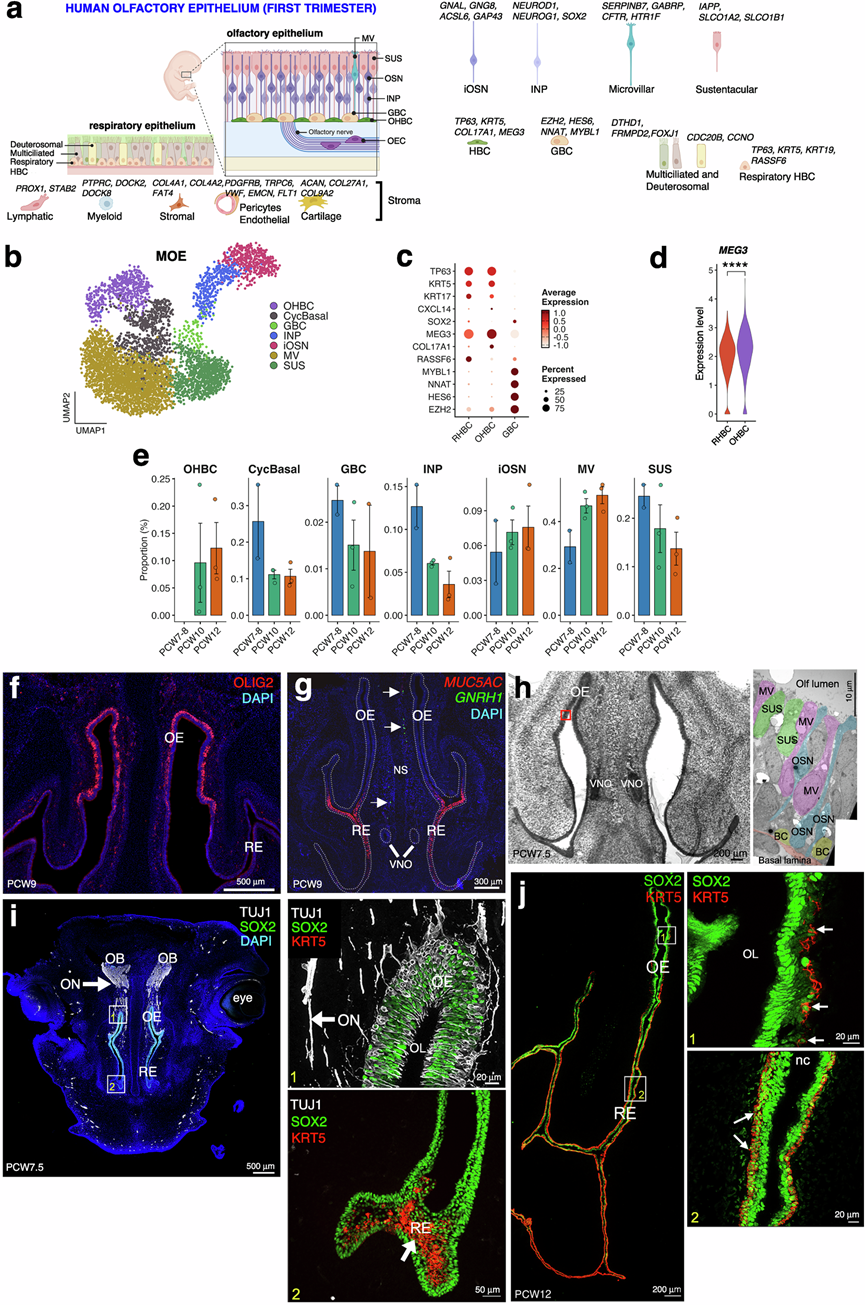
图2. PCW7-PCW12 处人类嗅觉系统的组成。
(a) 基于snRNA-seq数据重建的胎儿嗅觉和呼吸上皮及基质示意图,细胞类型由标记基因注释。(b) 主要嗅觉上皮的UMAP图。(c) 区分RHBC、OHBC和GBC的标记基因的散点图。(d) OHBC和RHBC中每个细胞MEG3表达的小提琴图 。(e) 各发育阶段每个样本的假性细胞类型比例。(f) 通过免疫荧光在 PCW9 时嗅觉和呼吸上皮冠状切片中验证标记物。(g) 在PCW9 时进行多重原位杂交以验证标记物。(h) 妊娠7.5周(PCW7.5)时嗅上皮(OE)的透射电镜图像显示了早期上皮细胞分化。(i) 妊娠7.5周(PCW7.5)时的免疫染色显示TUJ1+轴突从嗅上皮向嗅球方向延伸。(j) PCW12冠状鼻腔切片的代表性图像。
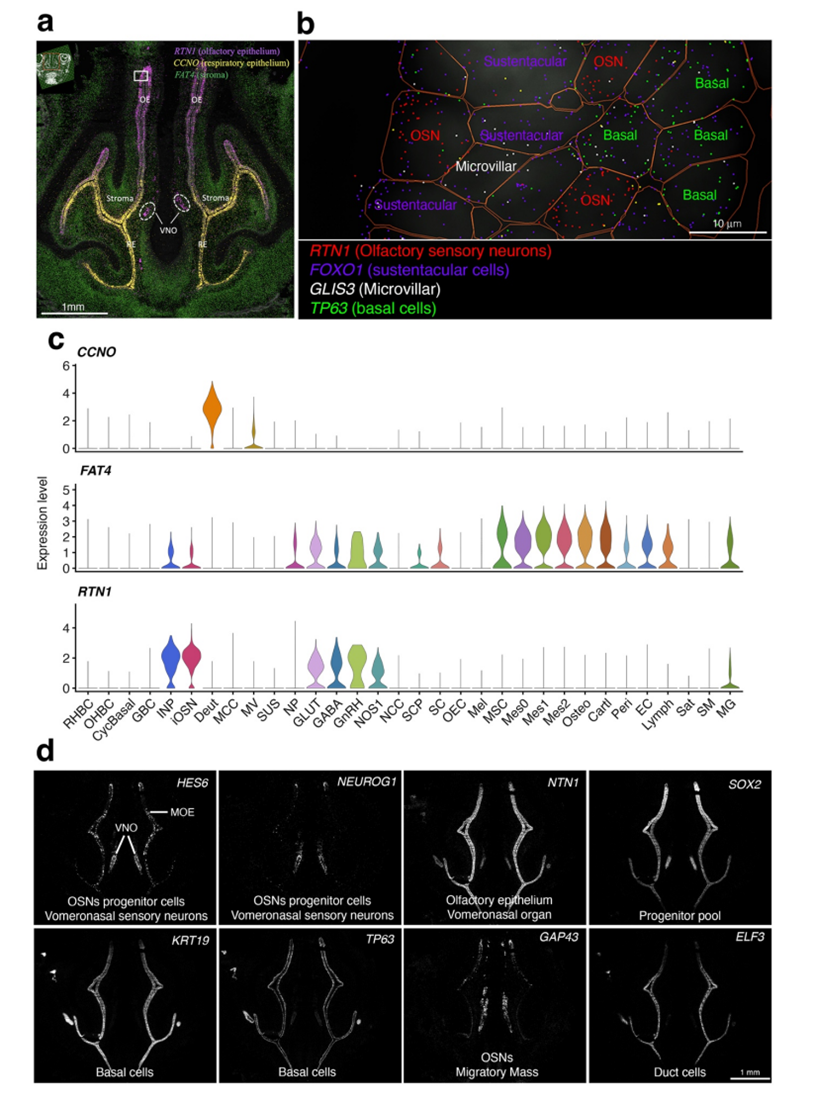
图S7. 发育中的嗅上皮的转录组学和空间谱。
(a) PCW9冠状鼻切面MERFISH图像显示RTN1、CCNO和FAT4表达。(b) 中框选区域的高倍放大图。(c) CCNO、FAT4 和 RTN1 在不同细胞类型中的表达小提琴图。(d) MERFISH 验证了定义嗅上皮细胞群的标记基因的表达情况。
03
人类胎儿嗅觉上皮的谱系推断和发育动态
鉴于先前记录的发育轨迹大多是在小鼠嗅觉上皮细胞中实现的,TM 可以探讨一些关于人类嗅觉上皮发育和胎儿干细胞微环境组成的问题。他们重点关注三种嗅觉细胞群:球状基底细胞(GBC)、中间神经元(INP)和嗅觉神经元(iOSN)(图 3a-b)。正如在成人人类嗅觉上皮中记录的那样,胎儿GBC层富含HES6和增殖标记物TOP2A的表达(图 3c)。其他在成人人类GBC中富集的经典标记物(NEUROG1和NEUROD1)在胎儿GBC中表达水平较低。相反,这些细胞富含与祖细胞增殖和早期神经元分化相关的转录因子,包括NNAT、MYBL1和FOXN4(图 3c)。
INP 的特征是TEX15、UNC5D、ANKFN1、FRMPD4的高表达水平,与神经元成熟和轴突靶向、神经元承诺和分化有关(图 3c)。本研究还发现,在人类胎儿嗅觉神经上皮中,GAP43、GNAL、ADCY3、PTPRR和SEMA3E在 iOSN 中表达(图 3c)。这些标记物在 iOSN 中的表达表明,神经元生长和嗅觉信号传导的关键成分在人类嗅觉神经元分化的早期阶段就已经参与其中。从 PCW7 到 PCW12 的时间序列分析显示,OE内嗅觉神经元的数量逐渐增加(图3d)。利用已知的标记基因对细胞类型进行鉴定,结果区分出 OHBC、CyclBasal、GBC、INP、iOSN、MV和SUS细胞(图 3e)。OSN的标记基因包括GNAL、GNG8、OLIG2和RTP1(图 3e)。
小鼠的谱系追踪表明,OE细胞起源于静止的HBC,经由增殖的GBC祖细胞分化而来,这些祖细胞可产生神经元和非神经元谱系。类似的谱系轨迹是否在人类中存在尚不清楚。为了解决这个问题,他们使用Slingshot技术对人类OE的snRNA-seq数据进行了轨迹推断,整合了聚类的身份信息和UMAP嵌入信息来推断分支谱系(图3f)。该分析解析出了起源于共同祖细胞的不同神经元、微绒毛和支持细胞的谱系轨迹,并得到了伪时间排序的支持(图3f-g)。伪时间密度分析显示,在早期发育阶段(PCW7-8),神经元和非神经元细胞均富集,随后在后期阶段(PCW10-12),细胞谱系逐渐向非神经元谱系(尤其是微绒毛谱系)转移(图 3h)。这些数据共同支持这样一种模型:GBC作为双潜能祖细胞,沿着神经元轨迹产生INP和iOSN,而其他分支则产生微绒毛谱系和支持细胞谱系(图 3i)。
动态基因表达热图揭示了沿推断轨迹的谱系特异性转录程序(图 3j)。神经元谱系表现出从祖细胞相关转录因子到神经元成熟基因的转变,而微绒毛谱系则表现出上皮分化和膜转运的早期激活,支持细胞则逐渐上调与解毒、结构完整性和信号传导相关的支持细胞程序(图 3j)。
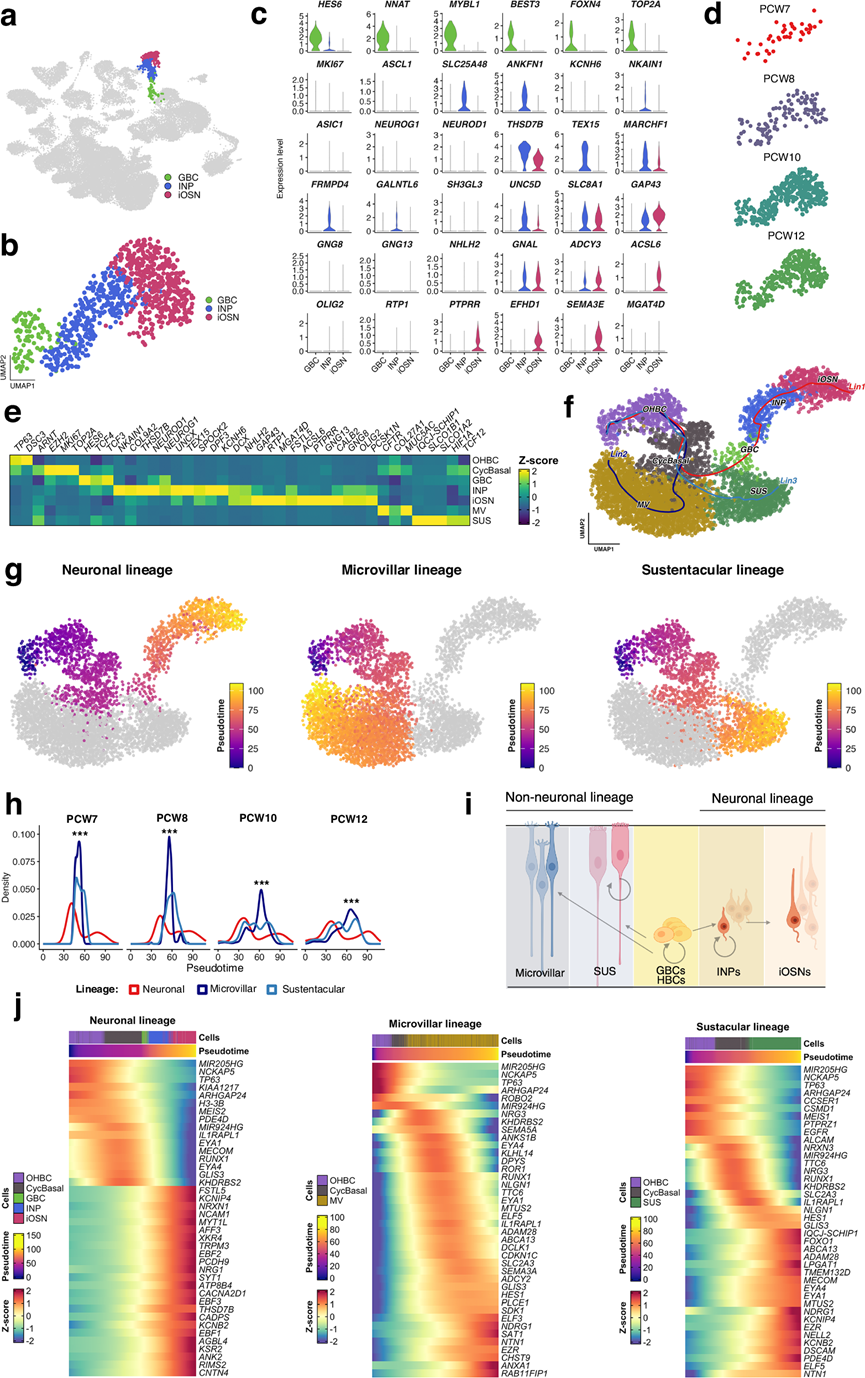
图3. 人类胎儿嗅觉上皮的谱系推断和发育动态。
(a) 嗅觉上皮细胞群的UMAP可视化。(b) 仅限于包含GBC、INP和iOSN细胞群的神经元谱系的UMAP子集。(c) 小提琴图显示了GBC、INP和iOSN细胞群中谱系特异性标记基因的对数归一化表达。(d) 根据受孕后周数(PCW7-PCW12)着色的嗅觉上皮细胞的UMAP投影。(e) 热图显示了选定标记基因在嗅觉上皮细胞群中的表达。(f) 基于Slingshot方法推断的人类胎儿嗅觉上皮发育轨迹。(g) UMAP 嵌入图,叠加了 Slingshot 推断的伪时间轨迹。(h) 密度图,展示了 Slingshot 推断的神经元、微绒毛和支持细胞谱系在不同发育阶段 (PCW7-PCW12) 的伪时间值分布。(i) 神经元(GBC → iOSN)和非神经元(GBC → MV)分化轨迹的示意图。(j) 热图,展示了神经元、微绒毛和支持细胞谱系在拟时间尺度上的基因表达动态变化。
04
转录调控因子和基因网络塑造人类嗅觉上皮发育
为了阐明发育中的人类OE中细胞命运决定的调控机制,他们推断了转录因子(TF)活性,并重建了不同细胞状态下的基因调控网络。TF调控子活性的比较揭示了不同的、谱系特异性的调控程序(图 4a)。静止的OHBC富集了KLF15、ZIC2和EGR2,而GBC和增殖期基底细胞则以增殖和祖细胞相关的TF为标志,包括E2F1/3/4、SOX2、ID1和ID3。相比之下,OSN显示出神经源性TF的激活,例如ASCL1、NEUROD1、NEUROG1、SOX4和ATOH1,这与神经元分化一致。这些调控程序也表现出发育阶段特异性,早期祖细胞(PCW7-8)与后期分化程度更高的细胞(PCW10-12)存在差异。
为了描述调控图谱,他们分析了所有细胞类型中的调控子活性(图 4b)。神经元群体表现出神经发育调控因子(包括PHOX2A、OTX2、NEUROD2、ARID1B、NFKBIZ、BARX2和HOXC8)的活性和表达升高,而这些调控因子在基底细胞和支持细胞中基本不活跃(图 4b)。这种组合表明,INP和iOSN受一个多层转录网络的调控,该网络整合了神经元分化、表观遗传重塑和环境响应。OSN中OR的表达依赖于与增强子元件(GI)的长程相互作用,在啮齿动物中,这些增强子元件由EBF1、LHX2和共激活因子LDB1组装成一个调控中心。单细胞研究表明,OSN祖细胞在成熟神经元中分化为单基因表达之前,会短暂地共表达多个OR。与此模型一致,他们在发育中的人类OE的INP iOSN中观察到EBF1活性增强,同时LHX2也表达增强(图 4a-b)。
为了整合这些发现,他们重建了OE中以TF为中心的调控网络(图 4c)。该网络揭示了围绕关键神经源性转录因子组织的高密度互连模块,这些模块主要调控与神经元分化、轴突导向和感觉功能相关的基因;而祖细胞和支持细胞相关的转录因子则调控与细胞周期控制、上皮组织和免疫相关通路相关的基因。
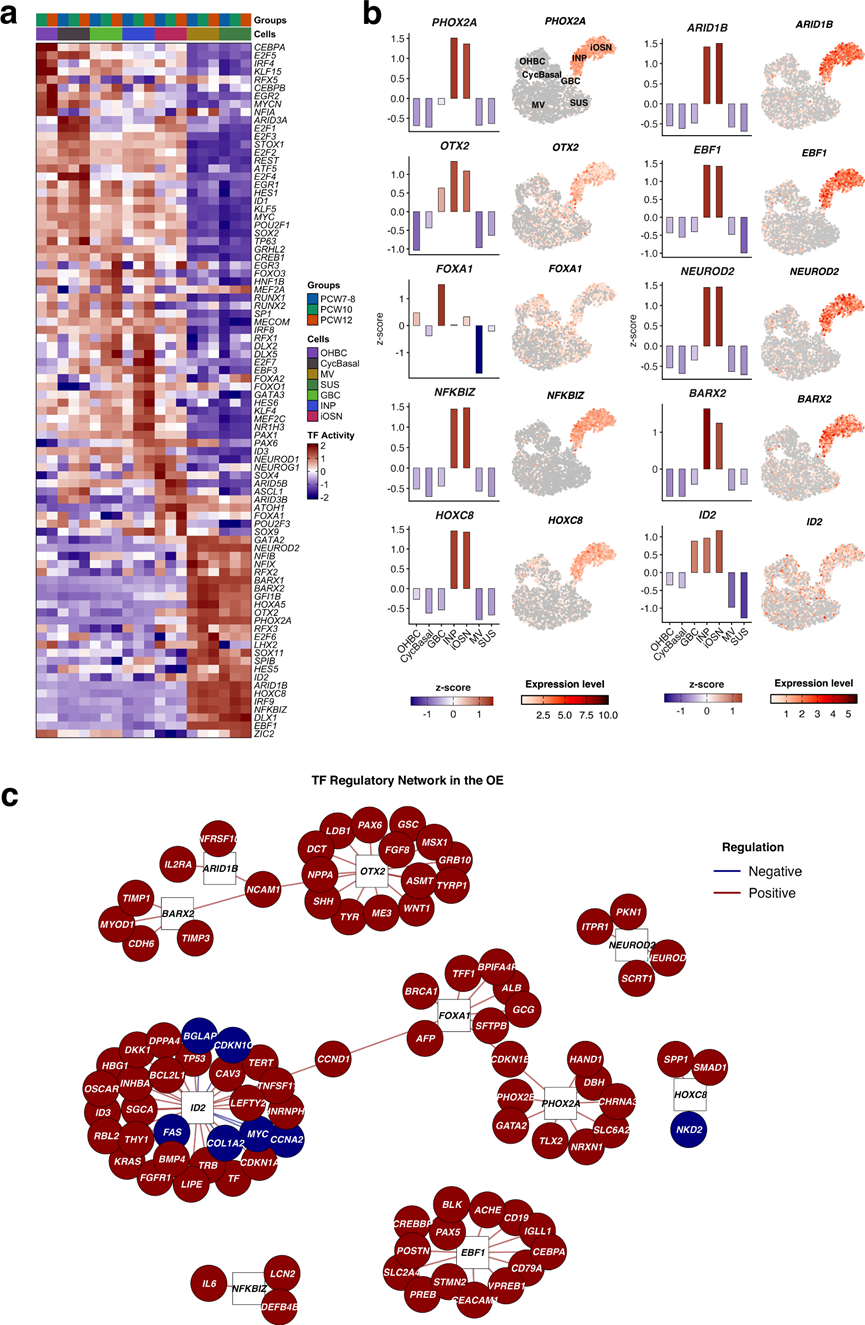
图4. 嗅觉感觉上皮中的转录因子(TF)活性。
(a) 人类嗅觉上皮(PCW7-12)发育阶段TF调控子活性的热图。(b) 条形图显示了不同嗅觉细胞类型中TF活性(左)及其对应的基因表达(右)。(c) 发育中嗅觉系统中TF相互作用的定向调控网络。
05
空间转录组学揭示了区域化的OE组织
接下来,他们利用空间转录组学解析了人类胎儿嗅觉上皮的空间组织和组织结构。为了将细胞群置于其天然微环境中进行分析,他们对人类胎儿头部冠状切片应用了MERFISH技术,分析了来自1例妊娠第9周胎儿(PCW9)的鼻上皮前部和后部区域,以及来自1例妊娠第 11周胎儿(PCW11)的后部切片(图 5a)。MERFISH技术能够对数百至数千个转录本进行高分辨率、单细胞水平的原位检测和定量。他们设计了一个包含294个基因的定制基因panel,其中包括通过snRNA-seq鉴定的细胞类型标记物和基于表达谱选择的57个OR转录本。
该分析生成了一个包含489030个细胞和58231407个检测到的转录本的数据集。将分析范围限定于解剖学定义的嗅觉上皮区域后,得到188803个细胞和16825698个转录本(平均每个细胞 89 个转录本)(图 5b-c)。为了实现跨样本的整合可视化,他们应用 Harmony 进行批次校正和降维,生成了空间转录组图谱的统一 UMAP 表示(图 5d)。
随后,他们使用scMusketeer方法将snRNA-seq数据集中的标签转移到MERFISH数据,从而实现了自动细胞类型注释。该方法利用了两种成像方式之间共有的294个基因表达谱。这种标签转移方法是稳健的,并且由此产生的空间细胞类型分配准确地重现了嗅觉上皮已知的分层结构和分区(图 5e)。包括PAX7、PAX6、LHX2和BMP家族成员在内的关键信号分子在嗅觉系统的模式形成、形态发生和神经元分化中发挥着核心作用,并引导嗅觉感觉神经元(OSN)轴突投射至嗅球。利用MERFISH技术,他们观察到,在人类胎儿组织中,与啮齿动物的研究结果一致,PAX7和BMP4划分了不同的间充质区域;PAX6在嗅觉和呼吸上皮中均有表达;而LHX2则选择性地富集于iOSN中(图5f)。这些空间表达模式与小鼠胚胎中报道的模式相似,支持了关键模式形成程序在不同物种间的保守性。
其他标记物,包括ELF3(呼吸道上皮导管细胞)、FOXO1(嗅觉祖细胞和基底细胞)和LHX2(iOSN),进一步界定了嗅觉上皮和呼吸道上皮之间的边界(图5g)。OR 基因的空间定位也强化了这种区分,这些基因仅限于嗅觉上皮(图 5h)。为了进一步解析嗅上皮的空间组织结构,他们比较了两个解剖学上不同的区域,即前部和后部的嗅上皮“脊”,将它们定义为从基底膜延伸至管腔并沿背腹轴分布的狭窄的、呈放射状排列的区域(图 5i-k)。聚焦于这些空间上连贯的单元,使他们能够评估前后方向和腹背方向的局部组织结构和细胞类型分层。与先前的人类研究一致,他们在PCW9处识别出了犁鼻器(VNO)及其相关的MM,主要位于最前部的切片中(图 5i)。在前部和后部区域,均可观察到清晰的腹背向RE到OE的过渡(图5j-k)。RE形成了一个富含多纤毛细胞、后体细胞和基底KRT+ 细胞的侧向区域,而OE则组织成嗅觉特异性层,包括OHBC、INP、iOSN和SUS(图 5j-k)。
虽然OE的前部和后部区域遵循相似的发育程序,但它们的上皮结构却表现出显著的区域差异。前部OE结构更为紧凑,OSN密集分布于背侧尖端,且OE与RE之间存在清晰的边界(图 5j)。相比之下,后部OE的嗅觉范围更广,RE-OE过渡更为平缓,OSN和SUS的分布也更为广泛(图 5k)。在两个区域中,INP和OSN均在背侧富集,这表明OE的扩张沿着腹侧到背侧的轴进行,导致上皮延长,同时也沿着内侧到外侧的轴进行,导致上皮增厚。
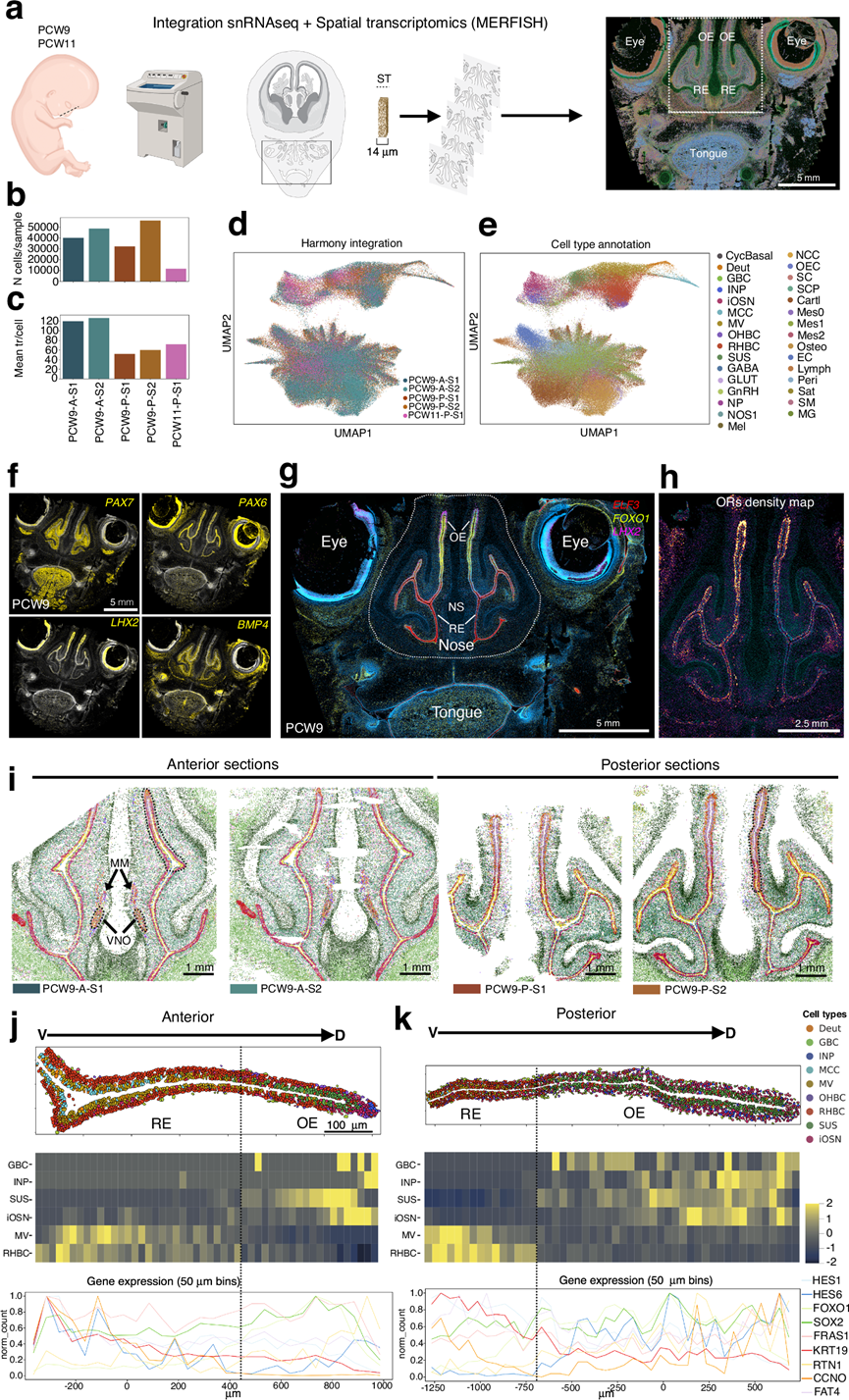
图5. 人类胎儿嗅觉发育的空间分辨单细胞图谱。
(a) 空间转录组学采样和工作流程示意图。(b) 每个样本的空间细胞总数。(c) 每个样本中每个细胞检测到的平均转录本数。(d) Harmony 整合 UMAP,按样本着色。(e) 细胞类型注释 UMAP,按细胞类型着色。(f) PCW9处覆盖鼻腔区域的冠状头部切片图像,使用PAX7、PAX6、LHX2和BMP4探针通过 MERFISH 成像。(g)使用不同的探针( ELF3、FOXO1、LHX2 )通过 MERFISH 成像。(h) 区域内嗅觉上皮(OE)中嗅觉受体密度图的放大图。(i) PCW9时期4个切片(前部2个切片,后部2个切片)的空间可视化图。(j-k) PCW9时期人类胎儿嗅觉上皮沿前后轴的细胞类型和基因表达的空间分布。
06
人类嗅觉上皮发育过程中嗅觉受体基因表达模式
对胎儿snRNA-seq数据集的转录组分析揭示了在产前发育的4个阶段,INP和iOSN中169个OR基因的表达(图 6a)。此外,他们还在单个OSN中检测到了犁鼻器1型受体1(VN1R1)的表达(图 6a)。在非神经元细胞群中,OR转录本几乎不存在。相反,OR表达在iOSN中显著富集,而在INP中则显著降低(图 6b)。他们随后评估了表达OR的INP和iOSN的比例(图 6c)。INP主要缺乏OR表达,而iOSN则表现出显著的OR阳性状态。他们观察到,超过40%的OR阳性iOSN仅表达单一受体,双表达的比例较低,表达三种或更多种OR的细胞则非常罕见(图6c)。这些数据表明,OR表达在从前体神经元到未成熟神经元的过渡期间开始,并迅速趋向于受限的受体使用,只有 10% 的 iOSN 表达 2 个 OR,而不到 1% 的 iOSN 表达 3 个或更多 OR(图 6c),这与人类产前发育过程中“神经元-受体法则”逐步建立相一致。他们既发现了 I 类 OR,也发现了 II 类 OR,其中 II 类 OR 占主导地位(图 6d),这与成人 OE的情况类似。此外,他们在一些非嗅觉细胞类型中也检测到了OR转录本。然而,与嗅觉上皮内的其他细胞群以及周围鼻腔区域相比,OR表达在iOSN中更为普遍(图 6e)。
接下来,他们基于基因组位置分析了OR的分布和表达(图 6f-g)。OR的数量和表达在基因组上的分布并不均匀;相反,OR基因座在1、7、11、12和19号染色体上更为密集(图 6f)。然而,这些基因座的转录活性并不一定更高(图 6g),这表明在每个基因组位点上,某些OR基因的选择机制已经局部激活,从而驱动特定基因的表达。为了评估单个OR的优势程度,他们基于表达量最高的OR与表达量第二高的OR的相对表达量计算了OR优势评分(图 6h)。优势评分低的细胞主要为非神经元细胞和INP细胞,而优势评分高的细胞几乎全部与iOSN细胞相关。iOSN细胞的比例随着优势评分的升高而逐渐增加,表明随着神经元成熟的进行,单个OR的优势地位不断增强。与此一致的是,高优势细胞的数量在发育过程中显著增加(图 6i)。在PCW7时,高优势细胞几乎不存在;在PCW8时,高优势细胞数量增加;到PCW10和PCW12时,高优势细胞数量显著增多,且主要集中在iOSN细胞群中。
随后,他们鉴定了在高显性细胞中最常检测到的显性OR基因(图 6j)。在PCW7和PCW8时,iOSN中检测到的显性OR基因很少,这与这些早期时间点OR表达的总体频率较低相一致。然而,到PCW10时,出现了一组清晰的显性OR基因,每个基因都由多个高显性细胞代表。到PCW12时,这一基因库进一步扩展,越来越多的OR基因在不同细胞中表现出重复的高显性表达。
最后,为了研究人类胎儿发育过程中OR的表达模式,他们分析了单个细胞(INP 和 iOSN)的 OR 转录情况。根据细胞的主要 OR 进行排序,他们观察到 OR 表达呈现出显著的对角线模式,表明单个神经元优先表达单个 OR 基因。值得注意的是,INP 细胞的 OR 表达有限,而强烈的表达则仅限于iOSN。主要OR表达的频率和强度从PCW7到PCW12逐渐增加,表明 OR 的选择在发育过程中逐渐稳定。总之,这些数据表明,OSN的标志性特征——单一 OR 表达——在人类胎儿发育早期就已经建立,并随着神经元的分化而不断完善。
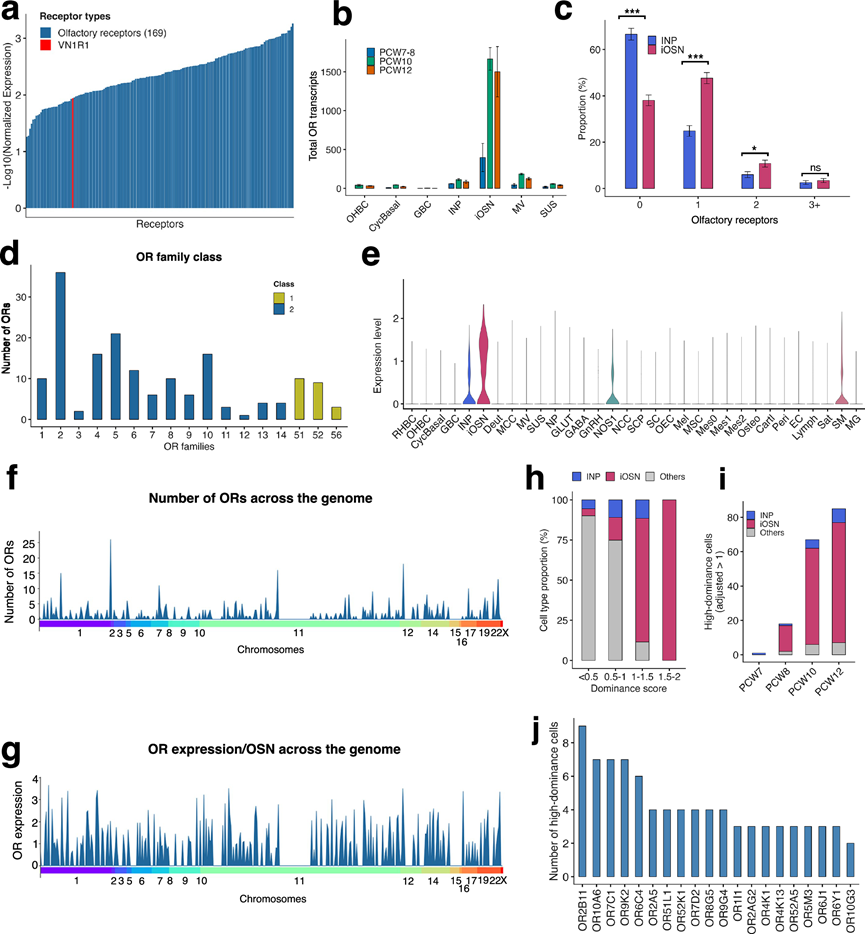
图6. 胎儿人类嗅觉系统中 OR 表达的分析。
(a) 在PCW7-12发育阶段,中间神经元祖细胞(INP)和未成熟嗅觉感觉神经元(iOSN)中检测到的嗅觉受体(OR)的表达水平。(b) 条形图展示了在PCW7-12发育阶段( n = 8个供体)构成嗅觉上皮(OE)的各种细胞类型中嗅觉受体(OR)转录本的总数,数值按供体汇总。(c) 样本中 INP和 iOSN的 OR 表达分布。(d) iOSN 和 INP 中各家族的 OR 数量。(e) 小提琴图显示胎儿鼻区嗅觉细胞和非嗅觉细胞中 OR 的分布。(f) OR 基因在染色体上的基因组分布。(g) 每个OSN 中 OR 表达水平的染色体定位。(h) 堆叠条形图显示按 OE 细胞类型分层的每个 OR 优势评分组别的细胞比例。(i) 按细胞类型分层,每个发育阶段的高优势细胞数。(j) 在高优势细胞中最常观察到的前 20 个优势 OR 基因。
07
空间分辨的嗅觉上皮OR基因谱分析
为了深入了解人类胎儿发育过程中OR基因表达的空间分布,他们应用MERFISH技术对OE内的OR转录本进行了高分辨率定位。MERFISH进一步揭示了PCW9和PCW11两个人类胎儿样本中嗅觉上皮内OR的表达情况。他们首先通过整合细胞间转录组相关性和物理邻近性来研究OE的空间组织,从而定义离散的空间区域(图 7a)。该分析揭示了一个高度特异性的区域D1016,对应于OE和RE。区域D1016由区域D1013包围,后者是一个异质性更高的区域,包含所有OE细胞类型以及OEC和雪旺细胞,提示其可能是一个支持性或过渡性微环境。其他空间区域也清晰地划分出来,并与已知的解剖学特征相对应:区域D1015包含软骨相关细胞,区域D1014富含成骨细胞,区域D1010对应于基质成分(图 7b)。
为了评估是否遵循“1个OR基因对应1个OSN”的规则,他们开发了一种调整后的优势度评分,其定义为每个细胞中表达量最高和次高的OR基因之间的表达差异,并以表达量最高的OR基因表达水平的对数进行分析。该指标同时考虑了优势度和表达强度,从而最大限度地减少了低表达转录本的干扰。将该评分应用于整个数据集,揭示了主要位于OE内的一组独特的、高分值的细胞(图 7c)。具有高优势度评分的细胞类型主要对应于iOSN,并且随着发育阶段的推进而增加,在PCW11达到最大值(图 7d),从而进一步证明了OSN中存在单一OR基因表达规则。
对不同空间位点的OR表达进行层次聚类分析,揭示了与特定细胞类型相关的OR基因使用模式(图 7e)。与其他嗅觉上皮细胞群相比,iOSN细胞中多种OR基因的表达显著富集,而非神经元细胞则几乎没有OR信号。他们利用MERFISH技术在PCW9和PCW11神经元上可视化了单个OR转录本以及典型的OSN标记物(图 7f)。在空间上离散的神经元聚类中检测到了OR基因,这些基因与OSN标记物(包括RTN1)共定位,这与OR基因的空间分离和早期选择相一致。在 PCW9 中,OR 表达稀少,绝大多数 OSN 表达 1 个单一的 OR,一些 OSN 共表达两个不同的 OR 基因,这与从转录组数据推断出的优势和共表达模式一致。在PCW11样本中,OR的表达更为普遍,在OE中检测到的OR基因数量和多样性均有所增加(图 7f)。值得注意的是,在这一后期阶段,单个OSN内多个OR的共表达现象很少见,这表明OR基因调控在发育过程中得到了精细化。
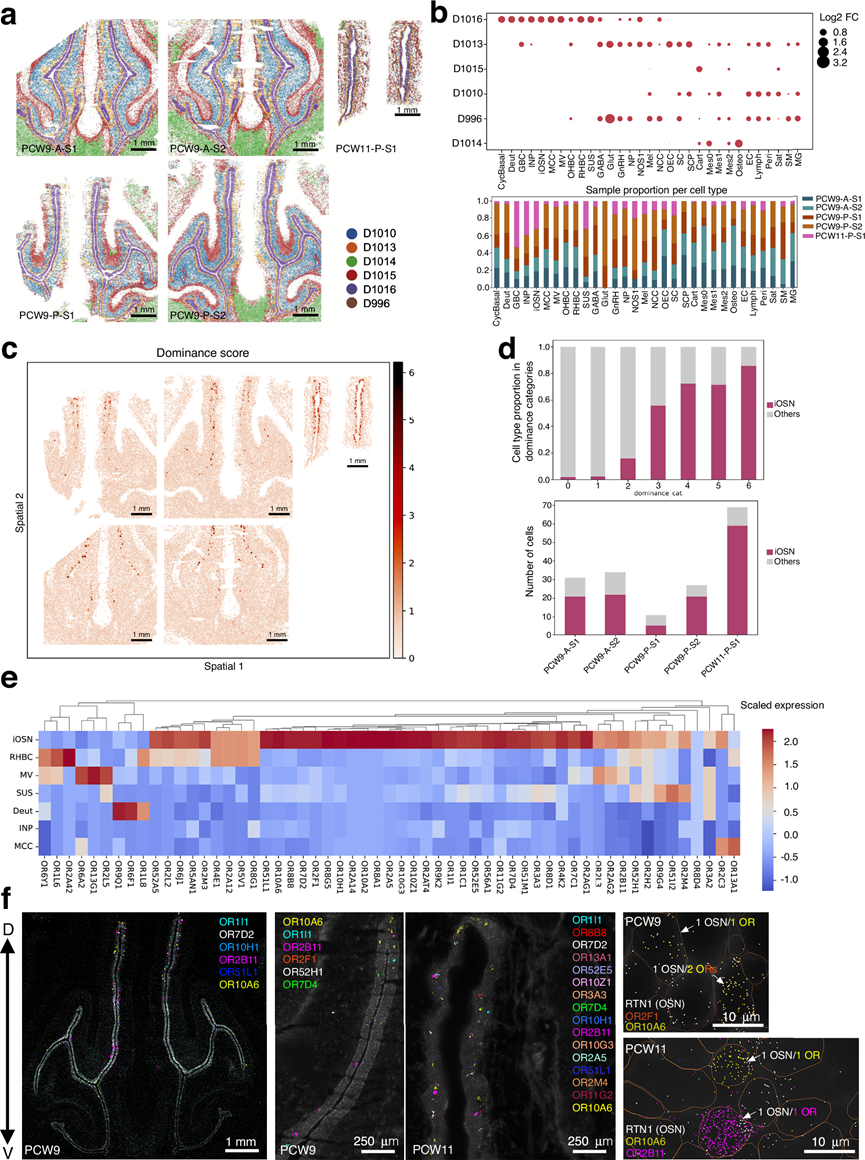
图7. MOE 中感觉神经元和嗅觉受体的空间分布。
(a) 跨切片的 Novae 空间域识别。(b) 上图显示 Novae 空间域内的细胞类型富集情况。(c) 比较每个细胞中最高嗅觉受体和第二高嗅觉受体的优势度得分。(d) 上图:优势度类别中的细胞类型比例。下图:嗅觉和呼吸上皮(Novae 域 D1013 + D1016)中优势度得分 >3 的每种细胞类型的细胞数量。(e) 热图显示代表性 OR 基因在不同上皮细胞类型中的表达量。(f) MERFISH 实验的代表性显微照片,显示 PCW9 和 PCW11 中列出的 OR 转录本的空间分布。
+ + + + + + + + + + +
结 论
本研究构建了人类胎儿鼻腔区域的snRNA-seq图谱,该图谱基于受孕后7至12周的男性和女性胎儿样本。snRNA-seq解析了32种不同的细胞类型,并结合MERFISH技术,实现了对OE及其邻近组织基因表达动态的时空定位。本研究鉴定了嗅觉感觉神经元分化的标志物以及调控上皮模式形成和OE形态发生的通路。值得注意的是,169个嗅觉受体基因的空间分辨snRNA-seq图谱揭示了“一个神经元对应一个受体”这一原则在妊娠早期就已存在的分子证据。这项工作共同建立了人类早期嗅觉发育的分子和空间框架,并为感觉神经发生和先天性疾病的研究提供了资源。
+ + + + +



