

 English
English文献解读|Cell Metab(30.9):肠道菌群通过重塑肠脑能量代谢来缓解抑郁症
✦ +
+
论文ID
原名:The gut microbiota alleviates depression by remodeling gut-brain energy metabolism
译名:肠道菌群通过重塑肠脑能量代谢来缓解抑郁症
期刊:Cell Metabolism
影响因子:30.9
发表时间:2026.03.31
DOI号:10.1016/j.cmet.2026.03.002
背 景
抑郁症是一种常见的精神障碍,影响着全球5%的人口,由于其高发病率和不尽如人意的治疗效果,造成了巨大的社会经济负担。值得注意的是,亚临床抑郁综合征的患病率甚至更高(17%),尤其是在儿童和青少年中。目前的治疗模式仍然不足,只有不到三分之一的患者接受了有效的干预。目前的抑郁症药物治疗靶向单胺通路,但存在起效缓慢、副作用多和复发率高等问题。像氯胺酮这样的速效药物也受到安全性和适用性问题的限制。这凸显了开发新型、更安全、更易获得的治疗方法的迫切需求。
实验设计
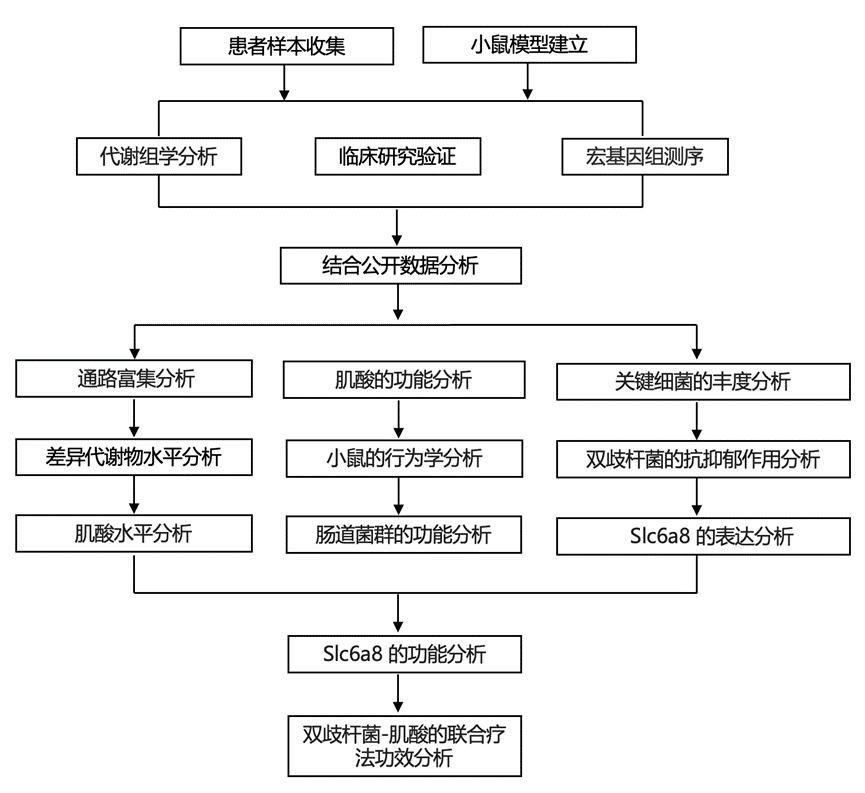
结 果
01

抑郁症患者的脑脊液、血浆和粪便代谢组学特征
为了研究抑郁症的代谢紊乱,研究团队对重度抑郁症(MDD)患者和健康对照组的脑脊液(CSF)、血浆和粪便样本进行了全面的代谢组学分析(图1A)。MDD患者和健康对照组均根据《精神疾病诊断与统计手册》(第五版;DSM-5)进行诊断,并使用DSM-5结构化临床访谈(SCID-5)和汉密尔顿抑郁量表(HAMD)-17进行评估。他们收集了41名受试者(15名MDD患者,26名健康对照者)的脑脊液样本进行非靶向代谢组学分析。主成分分析(PCA)显示MDD患者和健康对照者之间存在分离趋势。比较分析揭示了多种代谢物的差异表达,包括氨基酸、碳水化合物、维生素和肽类(图1B),KEGG通路富集分析显示甘氨酸、丝氨酸和苏氨酸代谢过程、乙醛酸和二羧酸代谢以及精氨酸和脯氨酸代谢通路富集(图1C)。血浆靶向代谢组学分析(31例重度抑郁症患者,22例对照)鉴定出12种上调代谢物和31种下调代谢物(图1D),其中精氨酸生物合成、丙氨酸-天冬氨酸-谷氨酸代谢和精氨酸-脯氨酸代谢均受到干扰(图1E)。此外,还收集了35名参与者(18例重度抑郁症患者和17例对照)的粪便样本。粪便代谢组学分析显示,37种代谢物上调,9种代谢物下调(图1F),这些代谢物富集于甘氨酸、丝氨酸和苏氨酸代谢,以及苯丙氨酸、酪氨酸和色氨酸的生物合成,以及精氨酸的生物合成(图1G)。对周围和中枢代谢改变的整合分析表明,精氨酸相关通路尤为重要(图1C-G)。值得注意的是,与这些通路相关的粪便代谢物在MDD患者中肌酸、谷氨酰胺、N-乙酰腐胺和天冬氨酸的水平显著升高(图1H)。虽然谷氨酸和天冬氨酸水平升高,但血浆和脑脊液中的肌酸水平降低(图1H-I)。这项综合代谢组学分析揭示了抑郁症中系统性肌酸失衡,其特征是粪便中肌酸水平升高,而血浆和脑脊液中肌酸浓度降低(图 1 I)。
为了进一步探究抑郁症动物模型中代谢物的变化是否与抑郁症患者的代谢物变化一致,他们采用了慢性社会挫败应激(CSDS)模型,这是一种广泛应用的抑郁症研究范式(图1J)。CSDS处理可诱导部分小鼠出现社交回避行为,这些小鼠称为应激易感(SUS)小鼠。对SUS小鼠和未受应激的对照(Ctrl)小鼠的粪便和血浆进行代谢组学分析,结果显示,通过主成分分析(PCA)可观察到明显的分离,并且KEGG通路分析显示精氨酸相关通路富集(图1K-L)。SUS小鼠与Ctrl小鼠粪便中富集于这些通路的差异代谢物表现为脯氨酸、4-胍基丁酸、谷氨酸、瓜氨酸和天冬氨酸的含量降低,而肌酸的含量升高(图1M)。有趣的是,虽然脯氨酸和天冬氨酸的减少在粪便和血浆中均一致,但SUS小鼠循环中的肌酸水平却降低,尽管粪便中肌酸积累(图1M)。SUS小鼠的脑脊液肌酸水平也降低(图1N)。鉴于先前有报道称,啮齿动物在应激状态下,其内侧前额叶皮层(mPFC)和海马中的肌酸水平会发生改变,他们接下来测量了小鼠在慢性社会应激(CSDS)后这两个脑区的肌酸浓度。与对照组(Ctrl)小鼠相比,SUS小鼠的mPFC肌酸水平显著降低(图1N),而海马肌酸水平保持不变(图1N)。不同生物隔室中肌酸水平的这种独特相反的趋势表明,抑郁症的病理生理机制中存在外周和脑部肌酸失衡。肌酸作为一种重要的能量缓冲剂,通过将高能磷酸基团从磷酸肌酸 (PCr) 转移到二磷酸腺苷 (ADP) 来再生三磷酸腺苷 (ATP)。越来越多的证据表明,肌酸具有改善抑郁症患者脑能量失衡的潜力。综上所述,基于跨物种趋同效应及其在抑郁症相关脑能量代谢中的既定作用,他们将重点研究肌酸的机制。
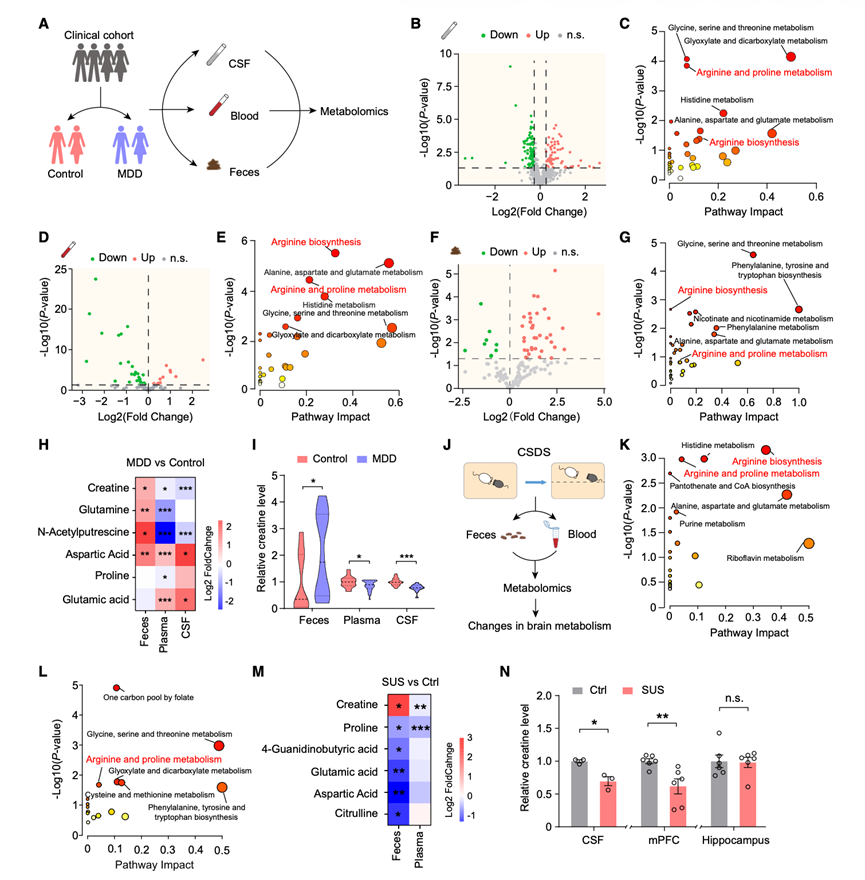
图1. 抑郁症患者的外周和脑代谢组学特征。
(A) 代谢谱分析实验方案。(B-C) 脑脊液中差异表达代谢物的火山图和 KEGG 通路富集。(D-E) 血浆中差异表达代谢物的火山图和 KEGG 通路富集。(F-G) 粪便中差异表达代谢物的火山图和 KEGG 通路富集。(H) 精氨酸相关途径代谢物的差异表达。(I) MDD 患者和健康对照者的粪便、血浆和脑脊液中的肌酸水平。(J-L) CSDS后Ctrl和SUS小鼠代谢谱实验概述。(M) SUS 小鼠与 Ctrl 小鼠中精氨酸相关通路代谢物的差异表达。(N) 肌酸水平。
02
外源性肌酸在小鼠中产生类似抗抑郁药的作用
肌酸是一种广泛使用的营养补充剂,膳食摄入量约能满足人体一半的肌酸需求。鉴于越来越多的证据表明肌酸对大脑健康有益,以及他们观察到的抑郁症患者肌酸失衡现象(图1),他们研究了外源性肌酸的潜在抗抑郁功效。成年雄性C57BL/6J小鼠在行为学测试前60分钟灌胃给予不同剂量的肌酸一水合物(0、10、100、500和1000 mg/kg)(图2A)。在强迫游泳试验(FST)(图2B)和悬尾试验(TST)(图2C)中,100、500和1000 mg/kg剂量的肌酸均显著降低了小鼠的静止不动时间。这些剂量在两种行为学测试中均表现出相似的疗效。在旷场实验(OFT)中,肌酸给药并未影响小鼠的运动活性。他们进一步评估了肌酸在慢性社会抑郁模型(CSDS)中的治疗潜力。在CSDS发生前14天及发生期间,他们给雄性小鼠饮用水中添加3 g/kg/天的肌酸一水合物或溶剂。该剂量由健康成年人的最大安全日摄入量转换而来。随后,他们评估了小鼠的社交回避行为和蔗糖偏好(图2 D)。暴露于CSDS的溶剂对照组小鼠的社交互动(SI)比率(图2 E-F)和蔗糖偏好(图2 G)均显著降低。值得注意的是,补充肌酸可有效预防这些CSDS诱导的缺陷,包括SI比率(图2 E-F)和蔗糖偏好(图2G)的降低。综上所述,这些结果表明外源性肌酸在小鼠中具有抗抑郁样作用。
为了确定肠道菌群是否是肌酸发挥抗抑郁作用所必需的,他们在给予肌酸前,先用抗生素(ABX)处理特定病原体清除(SPF)小鼠(图2H)。ABX处理后,小鼠的食物摄入量、体重或旷场试验(OFT)中的总体运动活性均未出现明显变化。虽然口服肌酸(100 mg/kg)显著降低了对照组小鼠在FST和TST中的不动时间(图2I-J),但这种作用在ABX处理的小鼠中消失(图2I-J),表明肠道菌群是外源性肌酸发挥抗抑郁样作用所必需的。
为了进一步研究抑郁症相关肠道菌群是否调节肌酸的抗抑郁样作用,他们对经ABX处理的SPF小鼠进行了粪便菌群移植(FMT),移植的粪便样本来自MDD患者(D-FMT)或健康对照组(H-FMT)(图2K)。小鼠口服肌酸一水合物(100 mg/kg),并在FMT后14天进行行为学评估(图2L)。与对照组相比,H-FMT小鼠在FST和TST中的不动时间显著减少,而D-FMT小鼠则无此现象(图2M-N),且两组小鼠的总体运动能力无差异。正常SPF小鼠口服肌酸后60分钟血浆肌酸水平迅速升高,而抗生素处理显著抑制了这一升高(图2O)。值得注意的是,与H-FMT小鼠相比,D-FMT小鼠的血浆肌酸水平也较低(图2O)。综上所述,这些结果表明,与抑郁症相关的肠道菌群介导了外源性肌酸的抗抑郁样作用。
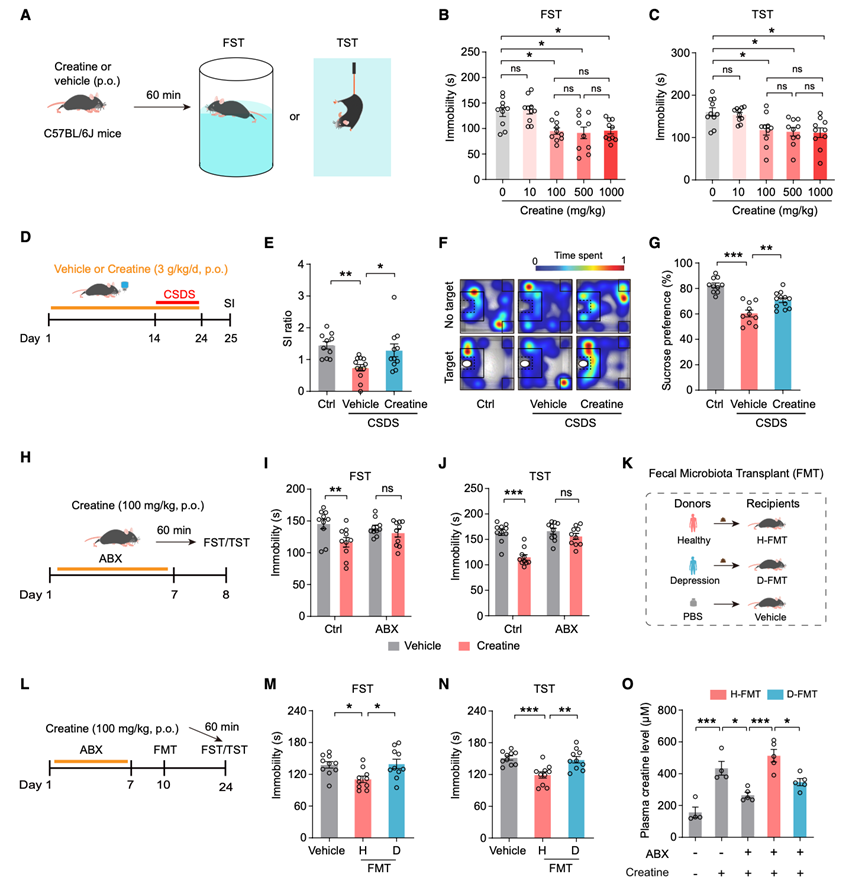
图2. 肠道菌群介导肌酸在小鼠体内的抗抑郁样作用。
(A) 实验设计示意图。(B-C) FST和 TST中的总不动时间。(D) 实验设计示意图。(E-F) SI 比率和 SI 测试中的代表性热图。(G) 小鼠对蔗糖的偏好。(H) 实验设计示意图。(I-J) 对照组和 ABX 处理组小鼠在 FST和 TST中的总不动时间。(K-L) FMT 研究的实验设计示意图。(M-N) 健康小鼠和抑郁症 FMT 小鼠在强迫游泳试验 (FST)和悬尾试验 (TST)中的总不动时间。(O) Ctrl、ABX、健康-FMT 和抑郁-FMT 小鼠的血浆肌酸水平。
03
双歧杆菌介导肌酸的抗抑郁作用
为了鉴定参与肌酸抗抑郁样作用的特定细菌类群,他们对MDD患者和健康对照组的肠道菌群组成进行了宏基因组测序。分类系统发育树显示,两组间双歧杆菌科的丰度存在显著差异,其中假长双歧杆菌(Bifidobacterium pseudolongum)在MDD患者中含量最低(图3A-B)。鉴于双歧杆菌的益生菌潜力,他们通过qPCR定量了几个菌种。MDD患者中假长双歧杆菌和青春双歧杆菌(B. adolescentis)的水平显著低于对照组,而长双歧杆菌(B. longum)和短双歧杆菌(B. breve)的水平则无变化(图3C)。同样,与H-FMT小鼠相比,D-FMT小鼠中假长双歧杆菌和青春双歧杆菌的水平也降低,且它们的丰度与血浆肌酸水平呈正相关(图3D-3F)。为了直接评估B. pseudolongum在肌酸吸收中的作用,研究人员将无菌(GF)小鼠接种双歧杆菌或PBS,然后给予氘标记的肌酸一水合物(肌酸-d5)。液相色谱-串联质谱(LC-MS/MS)分析显示,接种双歧杆菌的GF小鼠血浆中肌酸-d5水平显著高于PBS处理的对照组(图3G-H)。这些结果表明,双歧杆菌可能通过调节宿主肌酸平衡参与抑郁症的病理生理过程。
为了评估双歧杆菌是否介导肌酸的抗抑郁作用,他们用该菌定植了经ABX处理的小鼠。定植14天后,在口服肌酸一水合物60分钟后进行行为学测试(图3I)。定植了B. pseudolongum和B. adolescentis的小鼠在FST(图3 J-M)和TST(图3 K-N)中表现出显著减少的静止不动时间,而总体运动活性未发生改变。定植了假长双歧杆菌和青春双歧杆菌的小鼠血浆肌酸浓度升高(图3 L-O)。他们还用L. johnsonii作为对照菌株定植了经ABX处理的小鼠,结果显示其行为学未发生改变。口服肌酸一水合物后,感染L. johnsonii的小鼠在FST和TST中的不动时间未见显著变化,在OFT中的总体运动活性也未见显著变化。同样,两组小鼠的血浆肌酸水平也保持相当。
随后,他们研究了B. pseudolongum和肌酸联合给药是否能在抑郁模型中产生抗抑郁样作用。在CSDS模型中,SUS小鼠分别接受肌酸单药治疗或双歧杆菌联合肌酸治疗,疗程14天(图3P)。治疗前,与对照组相比,SUS小鼠口服肌酸60分钟后血浆肌酸水平显著降低。与单独使用肌酸相比,双歧杆菌联合肌酸治疗能更有效地改善SI比值(图3Q)和蔗糖偏好(图3R)。这种行为学改善伴随着血浆肌酸水平的显著升高(图3S)。综上所述,这些结果表明,B. pseudolongum和肌酸联合给药可在小鼠中产生显著的抗抑郁样作用。
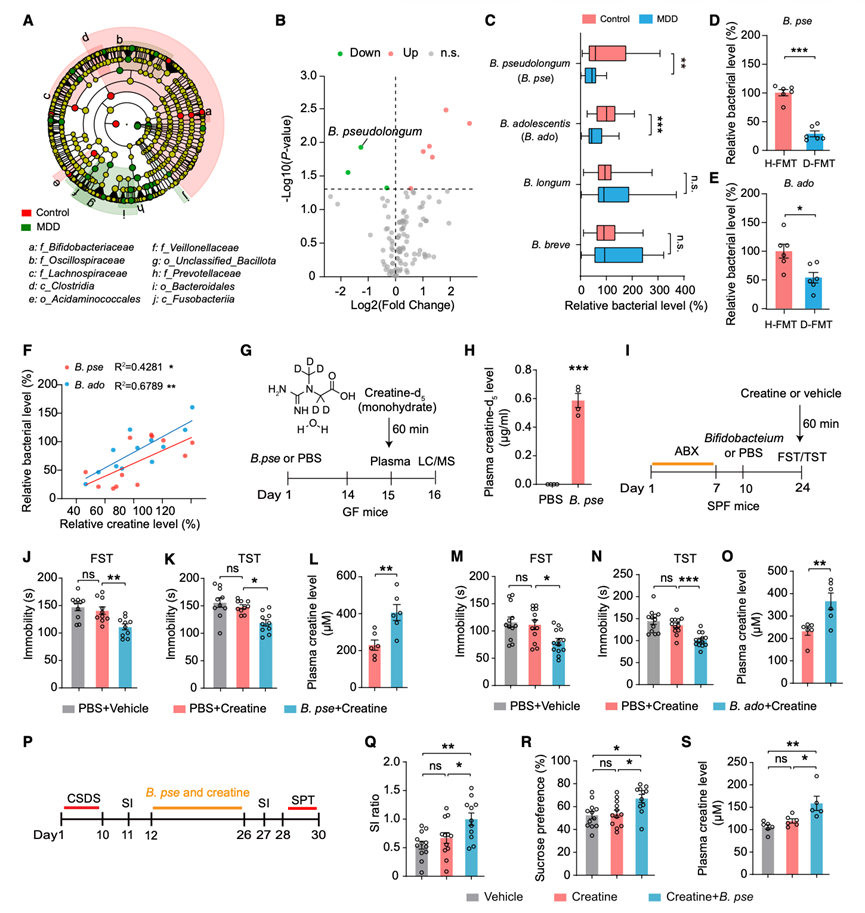
图3. 双歧杆菌介导肌酸的抗抑郁作用。
(A) MDD 患者和健康对照者粪便宏基因组学线性判别分析效应量(LEfSe)分析的分类分支图。(B) 双歧杆菌物种丰度火山图。(C) MDD 患者和健康对照组粪便中双歧杆菌的相对水平。(D-E) H-FMT 和 D-FMT 小鼠粪便中假长双歧杆菌和青春双歧杆菌的相对水平。(F) H-FMT 和 D-FMT 小鼠粪便中假长双歧杆菌/青春双歧杆菌水平与血浆肌酸水平的相关性。(G) 实验设计示意图。(H) GF小鼠血浆肌酸-d5水平。(I) 实验设计示意图。(J-L) 接种假长双歧杆菌或PBS的小鼠在强迫游泳试验(FST)、悬尾试验(TST)中的总不动时间,以及血浆肌酸水平。(M-O) 总不动时间以及接种B. adolescentis或 PBS 的小鼠的血浆肌酸水平。(P)实验设计示意图。(Q-R) SI 比率和蔗糖偏好。(S) SUS 小鼠血浆肌酸水平。
04
双歧杆菌产生的乙酸盐通过组蛋白乙酰化促进肠上皮细胞中Slc6a8的表达
由Slc6a8编码的肌酸转运蛋白(CrT)介导细胞对肌酸的摄取,对肠道肌酸吸收至关重要(图4A)。为了解双歧杆菌如何影响肌酸平衡,他们首先检测了Slc6a8的表达。在SUS小鼠的肠上皮细胞(IEC)中,Slc6a8的mRNA水平降低。与H-FMT小鼠相比,D-FMT小鼠的IEC中Slc6a8的mRNA水平也降低(图4B)。B. pseudolongum的定植增加了IECs中Slc6a8的mRNA水平。此外,假长双歧杆菌抑制了CSDS诱导的IECs中Slc6a8 mRNA和蛋白的下调(图4C-D),表明该细菌促进了Slc6a8的表达。
为了研究B. pseudolongum如何上调肠上皮细胞(IEC)中的Slc6a8表达,他们首先用假长双歧杆菌条件培养基处理肠道Caco-2细胞系。该处理显著提高了Slc6a8 mRNA的表达,表明细菌代谢产物可能是造成这种调控效应的原因。对条件培养基进行非靶向代谢组学分析,发现代谢组发生了显著变化,其中499种代谢物表达上调,327种代谢物表达下调(图4 E)。值得注意的是,与乙酸盐相关的代谢物是上调最显著的代谢物之一(图4 F)。乙酸盐检测证实,B. pseudolongum条件培养基(图4 G)和B. pseudolongum定植小鼠的小肠内容物(图4 H)中乙酸盐水平显著升高。鉴于短链脂肪酸(SCFA)乙酸盐可通过组蛋白乙酰化增加基因表达,在体内验证了这一机制(图4I)。给小鼠连续14天注射乙酸钠后,肠上皮细胞(IECs)中Slc6a8 mRNA和蛋白水平均显著升高(图4J-K)。染色质免疫沉淀测序(ChIP-seq)和ChIP-PCR实验证实,乙酸盐处理后,Slc6a8启动子区域的活性组蛋白标记H3K27ac富集增强(图4L-M)。
为了确定B. pseudolongum产生的乙酸盐对Slc6a8表达的必要性,他们构建了双歧杆菌的乙酸激酶(ackA)敲除菌株,因为ackA基因是双歧杆菌乙酸盐合成所必需的。与野生型(WT)双歧杆菌相比,ackA敲除双歧杆菌培养基中的乙酸盐水平显著降低。用ABX处理的小鼠,其肠道内容物中乙酸盐水平较低(图4 N-O),IEC中Slc6a8 mRNA(图4 P)和蛋白(图4 Q)水平也相应低于定植WT双歧杆菌的小鼠。总之,这些结果表明,双歧杆菌产生的乙酸盐通过增强组蛋白乙酰化促进肠上皮细胞中Slc6a8的表达。
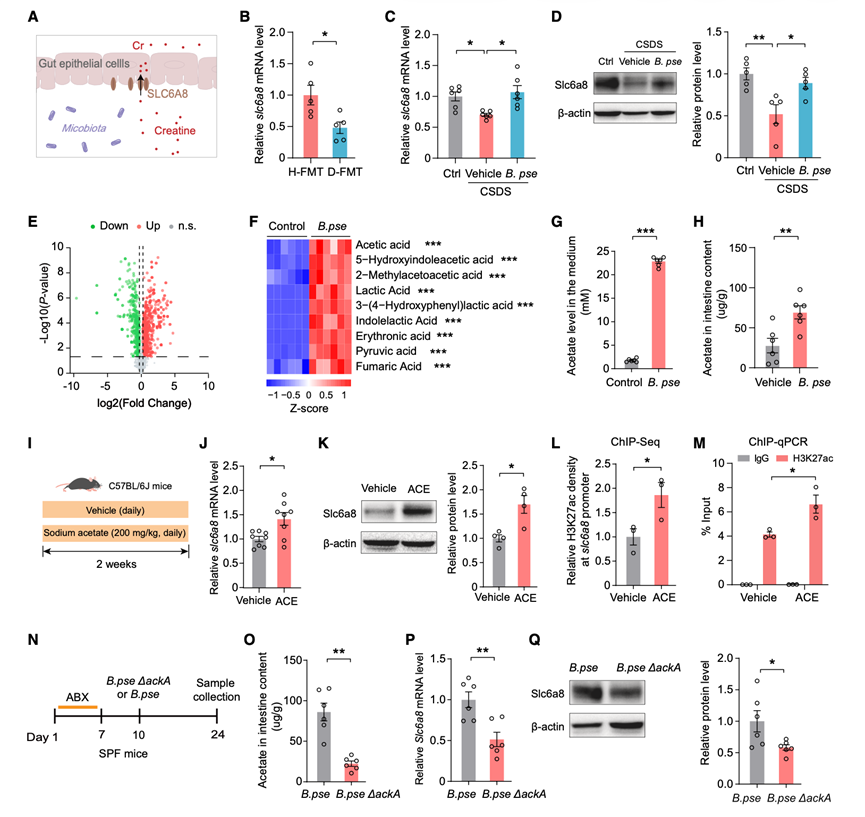
图4. 乙酸盐通过增强肠上皮细胞中组蛋白乙酰化促进Slc6a8的表达。
(A) 肌酸通过Slc6a8由IEC 吸收的示意图。(B) H-和D-FMT小鼠肠上皮细胞中Slc6a8 mRNA的相对水平。(C-D) Slc6a8 mRNA和蛋白质的相对水平。(E) 假长双歧杆菌条件培养基与对照培养基中代谢物的火山图。(F-G) 培养基中差异表达的代谢物和乙酸盐水平。(H) 接种假长双歧杆菌的小鼠小肠内容物中的乙酸盐水平。(I) 乙酸盐处理实验设计示意图。(J-K) 小鼠肠上皮细胞中Slc6a8 mRNA和蛋白质的相对水平。(L-M) ChIP-seq和 ChIP-PCR显示Slc6a8启动子区域的组蛋白乙酰化。(N)实验设计。(O) 用 WT 或ackA敲除B. pseudolongum定植的小鼠小肠内容物中的乙酸盐水平。(P-Q) 用 WT 或ackA敲除B. pseudolongum定植的小鼠肠上皮细胞中Slc6a8 mRNA和蛋白质的相对水平。
05
肠上皮细胞中的Slc6a8介导外源性肌酸的抗抑郁样作用
为了进一步证实Slc6a8在IEC中外源性肌酸抗抑郁样作用中的作用,他们通过将Vil1-cre小鼠与Slc6a8 loxp/loxp小鼠杂交,构建了IEC特异性Slc6a8敲除(VcKO)小鼠(图5A)。在雄性和雌性小鼠中,与未补充肌酸的对照组小鼠相比,VcKO小鼠在FST(图5B)或TST(图5C和S5E)中的不动时间以及血浆肌酸水平(图5D)均无显著差异。口服肌酸显著降低了对照组小鼠在FST和TST中的不动时间,而这种作用在VcKO小鼠中消失(图5B-C)。与VcKO小鼠相比,肌酸一水合物显著提高了对照组小鼠的血浆肌酸水平(图5D)。在OFT中,这些小鼠的总体运动活性未观察到显著差异。随后,他们在CSDS前14天和CSDS期间,分别给雄性小鼠注射肌酸一水合物或载体。之后,评估了小鼠的社交回避行为和蔗糖偏好(图5 E)。肌酸治疗显著抑制了CSDS诱导的对照组小鼠的SI比值和蔗糖偏好的降低,而IEC中Slc6a8的缺失则消除了这种效应(图5F-H)。这些结果表明,肠上皮细胞中的Slc6a8介导了肌酸的抗抑郁样作用。
最后,为了确定B. pseudolongum是否通过增加肌酸吸收来介导外源性肌酸的抗抑郁样作用,他们用双歧杆菌定植了经抗生素处理的对照组(Ctrl)小鼠和VcKO小鼠。定植后,小鼠分别口服肌酸或载体,随后进行行为学测试。在定植了双歧杆菌的对照组小鼠中,肌酸显著减少了FST和TST中的不动时间。而这种效应在定植了双歧杆菌的VcKO小鼠中消失。在OFT中,这些小鼠的总体运动活性未观察到显著差异。
肌酸转运蛋白(CrT)在构成血脑屏障(BBB)的微毛细血管内皮细胞(MEC)中表达,而血脑屏障是肌酸进入大脑的主要途径。此外,CrT在神经元中也高表达。为了研究脑内CrT在肌酸抗抑郁样作用中的作用,他们通过将Slc6a8小鼠与Cdh5-creER或CamkIIα-cre小鼠杂交,条件性地敲除了这些细胞类型中的Slc6a8基因,分别构建了内皮细胞特异性敲除(EcKO)和神经元特异性敲除(NcKO)小鼠(图5I-R)。EcKO和NcKO小鼠的内侧前额叶皮层(mPFC)中Slc6a8 mRNA和肌酸水平均降低。在雄性和雌性小鼠中,与对照组小鼠相比,EcKO 和 NcKO 小鼠在FST(图 5J-S)和TST(图 5K-T)中的不动时间均增加,而在OFT中,两组小鼠的总体运动能力无差异(图 5L-V)。口服肌酸并未显著降低 EcKO 和 NcKO 小鼠在 FST(图 5J-S)或 TST(图 5K-T)中的不动时间。随后,他们在CSDS前 14 天和 CSDS 期间,对雄性小鼠分别给予肌酸一水合物或载体(图5N-W)。肌酸治疗显著抑制了CSDS诱导的对照组小鼠的SI比值和蔗糖偏好性的降低,而内皮细胞和神经元中Slc6a8的缺失则消除了这种抑制作用(图5O-Z)。这些结果表明,内皮细胞和神经元中的Slc6a8介导了肌酸在小鼠体内的抗抑郁样作用。
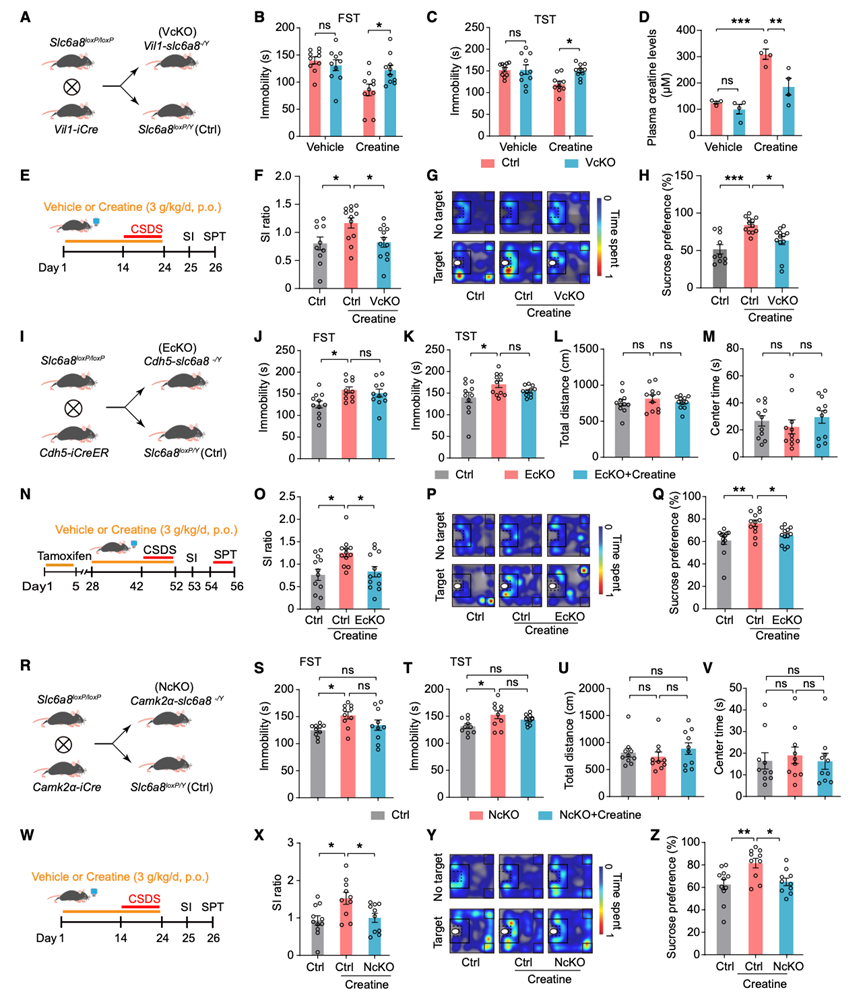
图5. Slc6a8介导肌酸的抗抑郁样作用。
(A) 针对IEC特异性Slc6a8敲除的遗传策略。 (B-C) Ctrl 和 VcKO 小鼠在 FST和 TST中的不动时间。(D) Ctrl 和 VcKO 小鼠的血浆肌酸水平。(E) 实验设计示意图。(F-H) SI 比率和代表性热图在 SI 测试和蔗糖偏好。(I) 内皮细胞特异性Slc6a8敲除的遗传策略。(J-M) FST 中的不动时间、TST 中的不动时间、OFT 中的总距离和中心停留时间。 (N) 实验设计示意图。(O-Q) SI 比率和代表性热图在 SI 测试中,以及 Ctrl 和 EcKO 小鼠的蔗糖偏好。(R) 锥体神经元特异性Slc6a8敲除的遗传策略。(S-V) Ctrl 和 NcKO 小鼠在 FST、TST中的不动时间,以及在 OFT 中的总距离和中心时间。(W) 实验设计示意图。(X-Z) SI 比率和代表性热图在 SI 测试和蔗糖偏好中 Ctrl 和 NcKO 小鼠。
06
神经元Slc6a8调节能量代谢和神经生理功能
鉴于肌酸在细胞能量代谢中已确立的作用,他们使用Seahorse XFe96分析仪对培养神经元中的线粒体功能进行了评估,包括线粒体应激测试和实时ATP速率测定(图6A)。他们用在突触蛋白启动子控制下表达Cre重组酶的慢病毒(LV-syn-cre)感染了源自Slc6a8条件性敲除小鼠的原代神经元,从而构建了条件性敲除神经元。Slc6a8的缺失显著损害了线粒体功能,降低了基础呼吸、ATP偶联呼吸和最大呼吸条件下的耗氧率(OCR)(图6B-C)。相应地,Slc6a8敲除神经元的线粒体ATP生成速率也显著降低(图6D)。这些结果表明,神经元中的Slc6a8影响能量代谢。
鉴于内侧前额叶皮层(mPFC)是抑郁症功能障碍的核心候选区域,且结果显示,在CSDS后,SUS小鼠mPFC中的肌酸水平(图1)和Slc6a8蛋白水平降低,他们研究了Slc6a8是否影响mPFC锥体神经元的神经生理功能。使用光纤光度计记录了NcKO小鼠和对照小鼠在强制互动测试(FIT)中,于亚阈值社会挫败应激(SSDS)前后,保护性金属网笼中的小鼠受到攻击小鼠攻击时mPFC锥体神经元的Ca2+活性(图6 E)。他们将AAV-CamkIIα-GCaMP6s病毒注射到NcKO小鼠和对照小鼠的mPFC中(图6 F),然后在感染细胞上方植入光纤以记录GCaMP荧光(图6 F)。在SSDS处理后,与对照组小鼠相比,应激的NcKO小鼠表现出更强的社交回避行为和更低的蔗糖偏好,表明SSDS诱导了NcKO小鼠出现抑郁样行为。在SSDS处理前,对照组和NcKO小鼠在攻击期的Ca2+信号均较非攻击期出现相似的升高(图6 G-I)。经过3天的SSDS处理后,发现NcKO小鼠在攻击期Ca2+信号的升高幅度较小(图6 G-I)。这些结果表明,神经元Slc6a8蛋白会影响对应激的Ca2+活性。
既往研究表明,在小鼠抑郁模型中,内侧前额叶皮层(mPFC)锥体神经元存在神经生理变化,包括Ca2+活性降低和微型兴奋性突触后电流(mEPSC)频率降低。激活mPFC锥体神经元可产生抗抑郁样作用。因此,他们采用离体全细胞膜片钳技术,测量了NcKO和Ctrl小鼠mPFC锥体神经元的兴奋性和mEPSC(图6J)。Slc6a8的缺失降低了神经元对逐渐增加的电流刺激的放电频率(图6K-L),并增加了阈值电流(图6M),但未改变静息膜电位、输入电阻或动作电位幅度。此外,NcKO小鼠的mEPSC频率显著降低,但幅度没有变化(图6N-P)。总之,这些结果表明神经元Slc6a8调节能量代谢和神经生理功能。
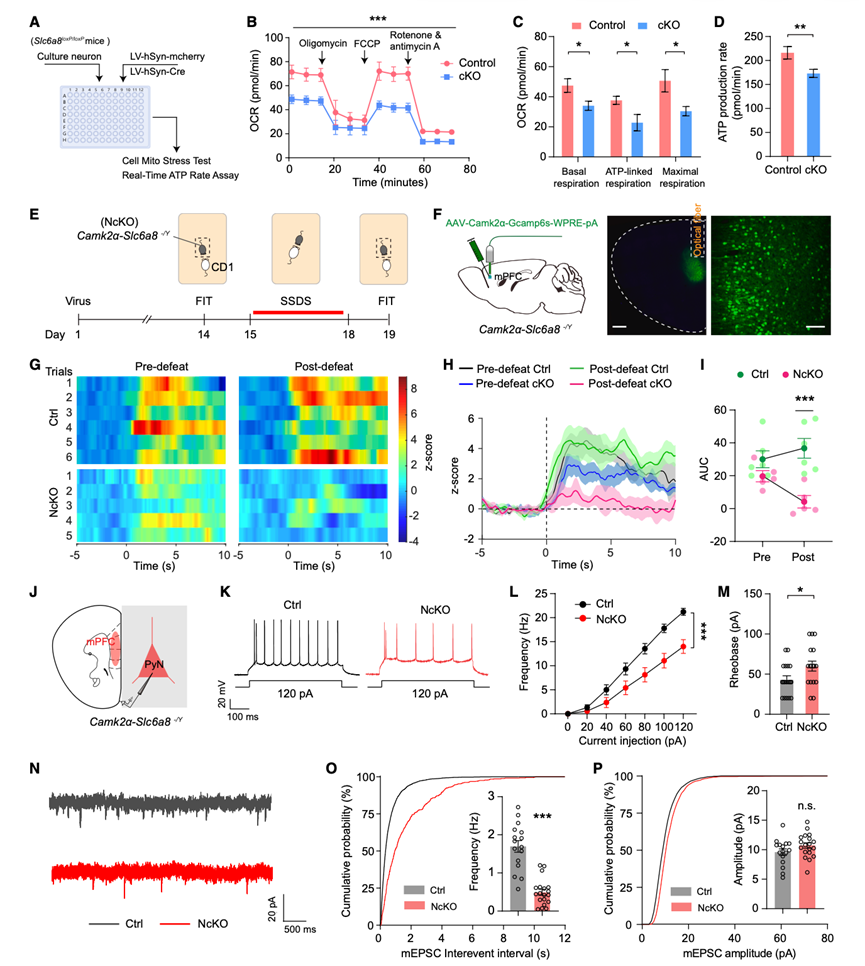
图6. 锥体神经元中的Slc6a8影响能量代谢和神经生理学。
(A) 实验设计。(B-C) 细胞线粒体应激测试中基线和应激条件下的 OCR 轨迹和定量。(D) 实时 ATP 速率测定中的线粒体 ATP 生成率。(E) 光纤光度测量的实验设计。(F) 纤维放置示意图和代表性的 GCaMP6 表达图像。(G) GCaMP6瞬态Z评分与社交互动事件相关的代表性热图。(H-I) 社交互动期间的平均Z分数变化和曲线下面积。(J) 实验设计示意图。(K) 由 +120 pA 的阶跃去极化电流引起的动作电位的代表性轨迹。(L-M) NcKO 和 Ctrl 小鼠动作电位的放电频率和阈值电流。(N) 来自 NcKO 和 Ctrl 小鼠的代表性 mEPSC 轨迹。(O-P) mEPSC 频率和振幅的累积分布和平均振幅。
07
双歧杆菌-肌酸组合对抑郁症患者的治疗效果
最后,他们开展了一项临床研究,以评估双歧杆菌联合肌酸补充剂对MDD患者的治疗潜力。首先,他们研究了在食品和医药领域已有应用的青春双歧杆菌是否能够提高健康成年人的血浆肌酸水平。16名健康受试者随机分为两组:一组单独服用一水肌酸(3 g/天),另一组同时服用青春双歧杆菌(2 × 108CFU /天)和一水肌酸(3 g/天),疗程为28天(图7A)。两组的基线特征和人口统计学数据具有可比性。基线时两组的血浆肌酸水平无显著差异,但在最后一次给药后1小时,所有受试者的血浆肌酸水平均显著升高。联合用药组的升高幅度比单独服用肌酸组更为显著(图7B),表明青春双歧杆菌能够提高人体血浆肌酸水平。
接下来,他们评估了青春双歧杆菌联合肌酸补充剂治疗抑郁症的潜在疗效。共有57名符合入组标准的MDD患者入组,其中45名患者随机分组。所有接受选择性5-羟色胺再摄取抑制剂(SSRI)治疗的MDD患者均接受青春双歧杆菌(2×108CFU/天)联合一水肌酸(3 g/天)或安慰剂治疗,疗程为28天(图7C)。安慰剂组22名患者中有19名完成了研究,青春双歧杆菌联合肌酸组23名患者中有19名完成了研究。完成研究的患者中,所有参与者均获得了抑郁评分,同时从9名安慰剂组患者和17名联合治疗组患者中采集了血液样本。两组患者的基线特征和人口统计学数据均衡良好。经过28天的干预后,联合治疗组的HAMD评分较基线显著降低,且改善程度优于安慰剂组(图7D-E)。同时,与安慰剂组相比,联合治疗组的血浆肌酸水平较基线显著升高(图7F-G)。这些结果表明,青春双歧杆菌联合肌酸治疗可能对抑郁症的治疗有益。
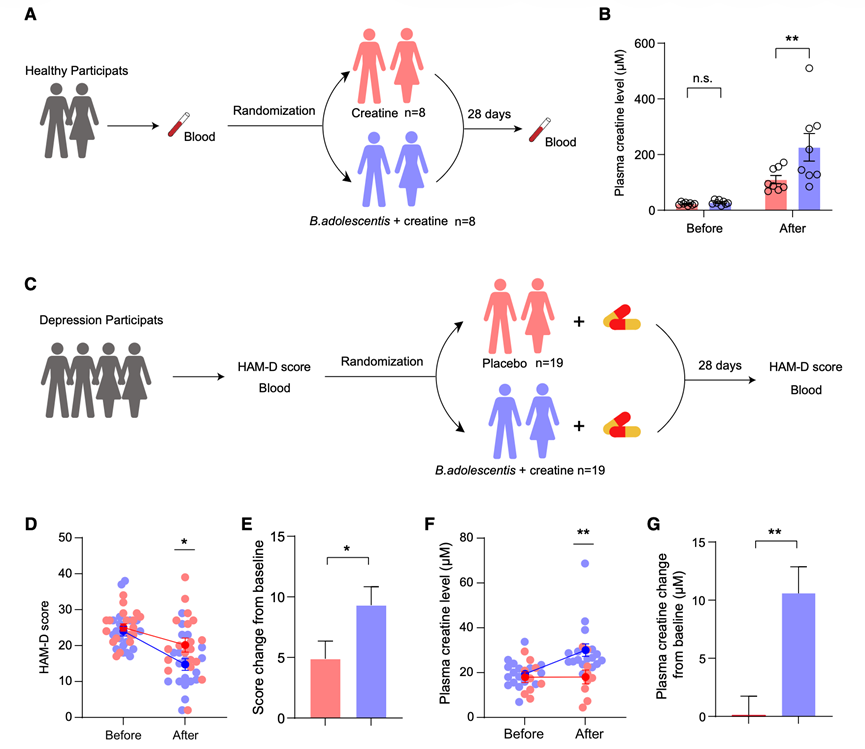
图7. 双歧杆菌和肌酸联合治疗MDD患者的疗效。
(A) 肌酸和青春双歧杆菌治疗健康人类的实验设计示意图。(B) 健康人体血浆肌酸水平。(C) 肌酸和青春双歧杆菌治疗 MDD 患者的实验设计示意图。(D-E) HAMD 评分(D)和 MDD 患者的基线变化。(F-G) MDD 患者的血浆肌酸水平及其与基线的变化。
+ + + + + + + + + + +
结 论
本研究利用代谢组学分析发现,抑郁症患者和抑郁症小鼠模型均存在粪便肌酸水平升高、血浆和脑脊液肌酸水平降低的情况。外源性肌酸通过肠道菌群发挥抗抑郁样作用。研究发现,抑郁症患者肠道中假长双歧杆菌的丰度显著降低,且与肌酸吸收受损相关。补充假长双歧杆菌可增强肌酸的抗抑郁作用。机制上,假长双歧杆菌产生的乙酸盐通过组蛋白乙酰化促进肠上皮细胞中Slc6a8的表达。Slc6a8介导了肌酸的抗抑郁样作用。神经元肌酸缺乏会影响能量代谢和神经生理功能。在服用抗抑郁药的抑郁症患者中,肌酸与双歧杆菌联合用药可提高血浆肌酸水平并降低抑郁评分。这些发现表明,双歧杆菌-肌酸组合是一种有前景的抗抑郁策略,并强调了肠脑能量代谢在抑郁症中的关键作用。
+ + + + +



