

 English
English文献解读|Cell Rep Med(10.6):黏膜相关淋巴组织淋巴瘤的跨组织图谱揭示了肿瘤内异质性和微环境亚型
✦ +
+
论文ID
原名:Cross-tissue atlas of mucosa-associated lymphoid tissue lymphomas reveals intratumoral heterogeneity and microenvironmental subtypes
译名:黏膜相关淋巴组织淋巴瘤的跨组织图谱揭示了肿瘤内异质性和微环境亚型
期刊:Cell Reports Medicine
影响因子:10.6
发表时间:2026.05.19
DOI号:10.1016/j.xcrm.2026.102785
背 景
黏膜相关淋巴组织(MALT)淋巴瘤是边缘区淋巴瘤中最常见的亚型,其特征是一组起源于不同黏膜部位的低级别结外B细胞恶性肿瘤。越来越多的证据表明,MALT淋巴瘤的发生涉及慢性抗原驱动的免疫失调,导致克隆扩增的B细胞发生恶性转化,特定的基因改变促进疾病进展。这种持续的抗原刺激可能源于慢性感染(例如,胃黏膜的幽门螺杆菌[HP]和眼附属器组织的鹦鹉热衣原体)或自身免疫性疾病(例如,干燥综合征和桥本甲状腺炎)。由于不同解剖部位的病因驱动因素各不相同,分子特征和淋巴瘤微环境 (LME) 存在显著的异质性,因此有必要进行进一步研究,以增进对机制的了解和精准医学方法。
实验设计
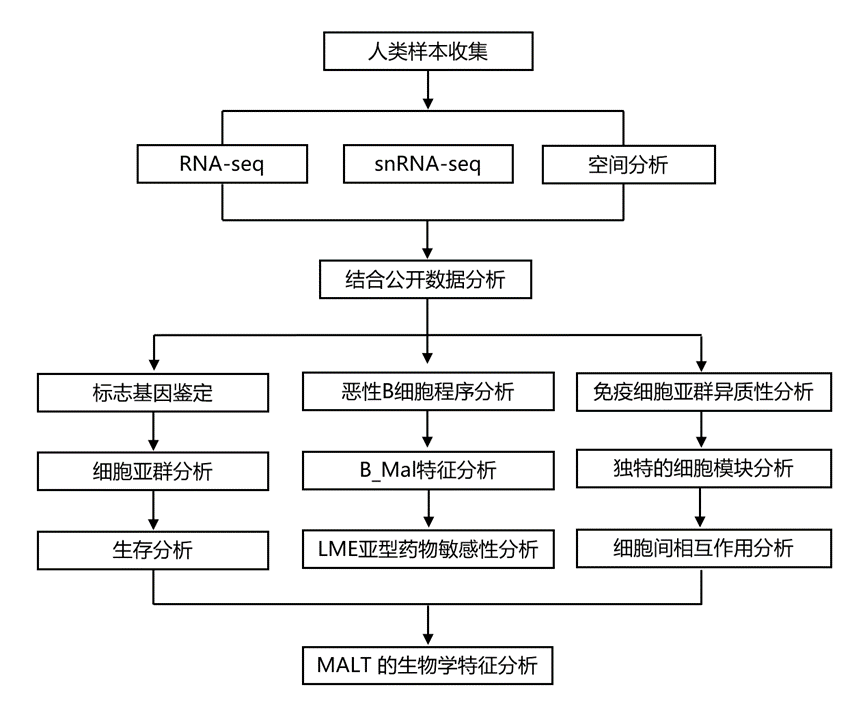
结 果
01

MALT淋巴瘤LME亚型的鉴定
研究团队回顾性收集了2020年1月至2023年6月期间,华西医院经病理诊断为MALT淋巴瘤的89例FFPE样本(WCH队列),其中大部分来源于胃(n = 38)和眼附属器(n = 30)。考虑到回顾性队列缺乏正常对照的局限性,他们进一步纳入了两个公开可用的包含眼附属器MALT淋巴瘤和正常组织的转录组分析(RNA-seq)队列(图1A)。最终,共有145个样本纳入整合分析,包括120个肿瘤样本和25个正常对照。在这些肿瘤样本中,89个来自内部的WCH队列,21个来自Shi等人报道的队列(GSE171059),10个来自Wolf等人报道的队列(GSE218618)。肿瘤样本主要来源于眼附属器和胃(图1B)。在120个肿瘤样本中,110个样本有临床信息。大多数患者为男性,年龄≥60岁,且在Ann Arbor分期为I期(图1C)。经批次效应校正和数据整合后,主成分分析(PCA)显示不同队列或肿瘤部位之间无明显分离。与正常对照组相比,MALT淋巴瘤样本中B细胞相关基因(如MS4A1、CD79A和PAX5)的表达显著上调,同时增殖相关通路(如E2F靶点和G2/M检查点)以及致癌通路(如未折叠蛋白反应和DNA修复)也富集。
为了解析120例MALT淋巴瘤样本中的LME亚型,他们对FFPE样本进行了单细胞核转录组分析(snRNA-seq)。考虑到样本质量和细胞核总数的技术要求,他们选择了16例手术切除样本进行后续的snRNA-seq分析(图1D)。去除背景噪声和进行质量控制后,他们对96778个细胞核进行了标准化、高变异基因选择、降维、批次校正和聚类分析。基于经典标记,他们注释了六种主要细胞谱系:B细胞、T细胞、髓系细胞、基质细胞、肝细胞和肺泡细胞,并使用均匀流形逼近和投影(UMAP)方法进行可视化(图1E)。
进一步的重新聚类分析在每个谱系内划分出多个亚群,这些亚群具有不同的标记表达谱(图1F)。为了区分恶性B细胞(B_Mal)和正常B细胞(B_Nor),他们应用inferCNV对拷贝数变异(CNV)进行评分,并应用基因集变异分析(GSVA)对边缘区B细胞(MZB)特征进行评分。B_Mal的CNV和MZB评分均显著高于正常B细胞,这与MALT淋巴瘤的MZB起源一致。B_Mal占B细胞的绝大多数,这与先前对脾脏边缘区淋巴瘤的单细胞研究结果一致。
为了最大限度地减少邻近正常组织的污染,他们评估了不同解剖来源的香农熵。五个熵值较低的亚群,每个亚群在肺或肝脏样本中均表现出显著富集(>80%),鉴定为可能来源于FFPE标本中的正常组织,因此排除在后续分析之外。接下来,他们基于亚群构建了MALT淋巴瘤特异性微环境特征,并应用GSVA进行特征评分,随后进行非负矩阵分解(NMF)聚类。该方法识别出三种不同的LME亚型:LME1(间质型),其特征是基质成分富集;LME2(炎症型),以淋巴细胞富集为特征;以及LME3(耗竭型),表现出整体微环境特征减弱(图1 G)。重要的是,这些LME亚型之间的差异与使用先前报道的LME相关基因集进行GSVA分析的结果高度一致。
根据LME分类,16个snRNA-seq样本包括4个LME1样本、8个LME2样本和4个LME3样本。对snRNA-seq数据中剩余亚群比例组成的PCA分析显示,不同LME亚型之间存在部分一致的模式。优势比分析表明,LME1富集内皮细胞、周细胞、中性粒细胞和Mac_MARCO,而LME2富集T细胞、树突状细胞(DC)和Mac_SPARC,这与基于bulk RNA-seq的LME分类结果一致(图1 H)。
与临床特征的关联分析表明,胃黏膜相关淋巴组织淋巴瘤(MALT淋巴瘤)和BIRC3/MALT1融合在LME1亚型中更为常见(图1I)。TNFAIP3失活突变的频率在LME亚型间无显著差异。生存分析显示,LME2亚型、晚期以及BIRC3/MALT1融合与更差的无进展生存期(PFS)显著相关(图1J)。此外,这三个因素在单因素和多因素Cox回归分析中均具有显著性,表明它们是MALT淋巴瘤患者PFS的独立预后因素。
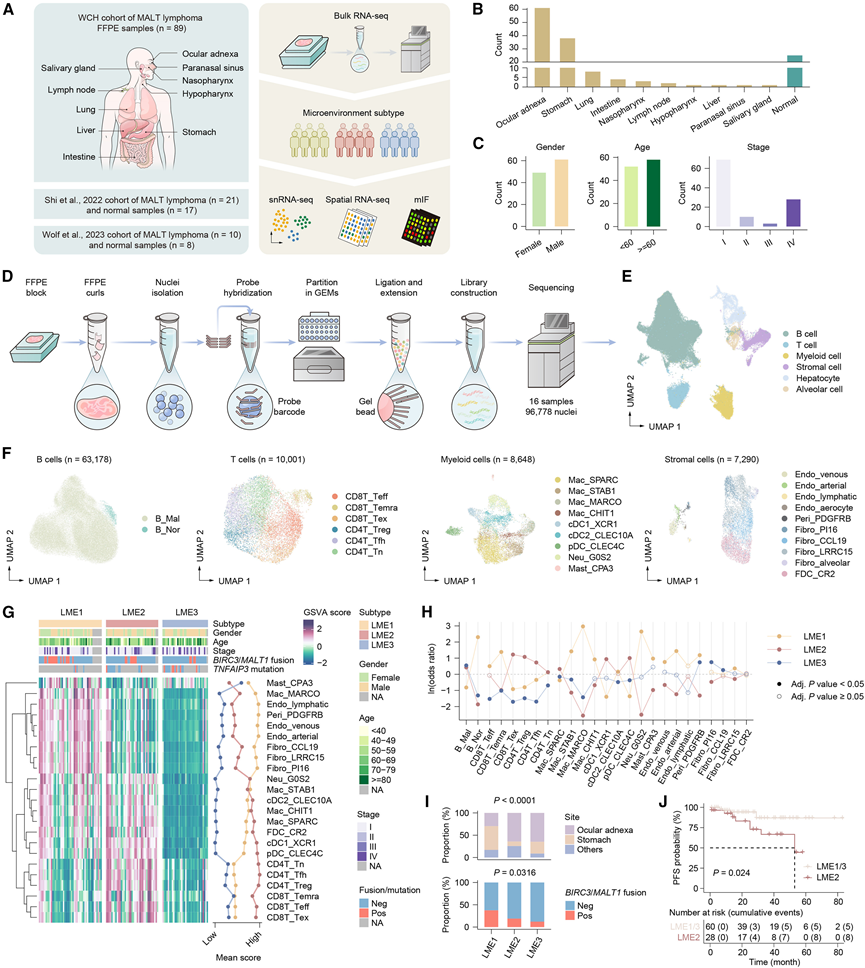
图1. MALT 淋巴瘤 LME 亚型的鉴定。
(A) 研究设计和工作流程示意图。(B) 柱状图显示了整合的批量 RNA-seq 数据集中来自不同解剖部位的 MALT 淋巴瘤样本和正常对照的数量。(C) 柱状图显示样本按临床特征分布情况。(D) 测序示意图。(E) 通过 snRNA-seq 分析的 96,778 个细胞核的 UMAP 图,描绘了六个主要细胞谱系。(F) 通过 snRNA-seq 分析的 63176 个 B 细胞、10001 个 T 细胞、8648 个髓系细胞和 7290 个基质细胞的 UMAP 图。(G) 热图(左)展示了120例MALT淋巴瘤样本中细胞亚群特征和临床特征的活性评分,并标示了三种LME亚型。折线图(右)显示了每种特征在三种LME亚型中的平均评分。(H) 折线图显示不同LME亚型中细胞亚群的富集情况。(I) 不同LME亚型中解剖部位(上图)和BIRC3 / MALT1融合(下图)的相对比例。(J) 根据LME亚型对具有随访数据的整合队列进行分层,绘制PFS的Kaplan-Meier曲线。
02
恶性B细胞的功能异质性和治疗脆弱性
与正常B细胞相比,恶性B细胞表现出细胞增殖和B细胞受体信号通路的显著上调。基于先前已建立的肿瘤内异质性分析方法,他们使用非负矩阵分解(NMF)分析了恶性B细胞的异质性,将其表达谱分解为不同的基因表达程序。在聚类参数变化下保持稳定且在多个样本中共有的程序定义为稳健程序。这些稳健程序基于共有基因的比例进一步聚类,最终得到7个元程序(MP),每个元程序包含50个代表性基因(图2 A)。
功能注释将这些MP与不同的过程联系起来,包括细胞周期G1/S期(MP1)、未折叠蛋白反应(MP2)、细胞周期G2/M期(MP3)、B细胞活化(MP4)、核酸结合(MP5)、免疫球蛋白生成(MP6)和免疫调节(MP7)(图2A-B)。在弥漫性大B细胞淋巴瘤的单细胞研究中也发现了细胞周期和核酸结合特征,表明B细胞淋巴瘤亚型之间存在共同的恶性B细胞程序。值得注意的是,MP6反映了与MALT淋巴瘤中浆细胞分化一致的免疫球蛋白程序,并且其评分与增殖性MP3和CytoTRACE2推断的发育潜能均呈负相关,支持终末分化恶性B细胞状态(图2C-D)。
他们从snRNA-seq定义的恶性B细胞中提取出B_Mal特征,例如经典的B细胞标志物CD79A和MS4A1,以及在淋巴瘤中高表达的CD37。将B_Mal特征应用于RNA-seq样本进行评分,并根据中位评分将患者分为两组(图2E)。单变量Cox回归进一步确定MP1、MP3和MP7为不良预后因素(图2F)。MP1和MP3的不良关联可能反映了它们与增殖活性增强的联系,而MP7包含趋化因子CCL22和CCL17,已知这些趋化因子可募集调节性T细胞(Treg)。与此一致的是,MP7评分与CD4+ T细胞中CD4+T_Treg细胞的比例呈正相关(图2G)。
有趣的是,尽管批量转录组数据受到非恶性谱系的影响,但来自 snRNA-seq 和 bulk RNA-seq 的 MP 评分在 LME 亚型之间基本一致。MP 评分并未因临床特征(如分期和年龄)而出现显著分层,这表明 LME 分类能更好地反映恶性 B 细胞的功能状态(图 2H)。例如,LME2 显示出较高的 MP4(B 细胞活化)和 MP7(免疫调节)评分,而 LME3 则富集了 MP3(细胞周期 G2/M 期)。使用 Seurat 计算的细胞周期评分也支持 LME3 具有更高的 G2/M 期评分和 G2/M 期细胞周期比例。
利用 Beyondcell 进行的药物敏感性预测进一步揭示了不同的治疗脆弱性:LME1 对血管内皮生长因子受体 (VEGFR) 抑制剂(索拉非尼、阿昔替尼和舒尼替尼)更敏感,这与其富含内皮细胞的微环境相符;LME2 对布鲁顿酪氨酸激酶 (BTK) 抑制剂伊布替尼更敏感,这与其较高的 MP4 评分相符;LME3 对长春新碱和长春瑞滨等可导致 G2/M 期阻滞的微管靶向药物敏感,这与其 MP3 富集相符(图2I)。基于靶点的药物敏感性预测(使用 scRANK)进一步证实了这些结果,表明 LME1 样本对 VEGFR2 靶向药物更敏感,而 LME2 样本对 BTK 靶向疗法更敏感。免疫组织化学分析提供了进一步的验证,结果显示 LME1 和 LME2 样本中 VEGFR2 和 BTK 的表达分别较高,而 Ki-67 表达的差异进一步证实了 LME3 中 G2/M 期细胞比例较高。这些亚型特异性反应特征凸显了 LME 分类在治疗分层方面的潜在临床应用价值。
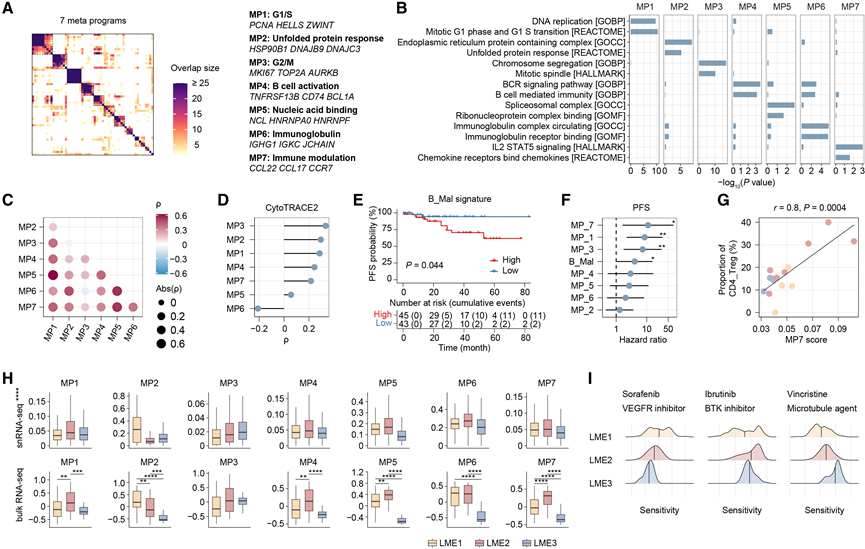
图2. 恶性 B 细胞的功能异质性和治疗脆弱性。
(A) 热图显示了基于前 50 个基因的稳健 NMF 程序之间的相似性。(B) 条形图显示通过对每个 MP 的基因集进行过度表达分析而确定的富集通路。(C) B_Mal 中 MP 特征得分之间的成对相关性。(D) CytoTRACE2评分与B_Mal中每个MP的特征评分之间的相关性。(E) 根据B_Mal特征评分对具有随访数据的整合队列进行分层,绘制PFS的Kaplan-Meier曲线。(F) 森林图描绘了按各MP和B_Mal特征评分分层的PFS风险比。(G) MP7 特征评分与 CD4 + T 细胞中 CD4_Treg 比例之间的相关性。(H) 箱线图展示了snRNA-seq样本(上图)和RNA-seq样本(下图)中各MP在不同LME亚型中的特征得分。(I) LME亚型药物敏感性分布的密度曲线。
03
LME亚型中免疫细胞亚群的多样性
为了阐明LME亚型间的免疫多样性,他们将主要免疫谱系进一步细分为更精细的亚型。T细胞分为CD8+亚群(CD8T_Teff、CD8T_Temra和CD8T_Tex)和CD4+亚群(CD4T_Treg、CD4T_Tfh和CD4T_Tn)。亚群比例因LME而异:CD8T_Tex在LME2中的比例显著高于LME1,而CD8T_Temra则呈现相反的模式(图3A)。根据经典标志物的表达,他们将CD8+ T细胞亚群分为三组:CD8T_Teff,其CCL5和GZMK高表达,对应于效应记忆表型;CD8T_Temra,其GNLY、GZMB和PRF1高表达,对应于细胞毒性和终末效应记忆T细胞(Temra); CD8T_Tex 由HAVCR2、LAG3和PDCD1定义,对应于耗竭表型。对于 CD4+T 细胞,CD4T_Treg 表达FOXP3以及IL2RA和TNFRSF4,对应于 Treg 细胞;CD4T_Tfh 表达CXCL13以及PDCD1和CXCR5,对应于滤泡辅助性 T (Tfh) 细胞;CD4T_Tn 表达IL7R以及CCR7和SELL,对应于初始 T 细胞(图 3B)。这些基于标记的分类得到了功能基因集富集评分的证实;例如,CD8T_Temra 的细胞毒性评分最高,而 CD8T_Tex 的耗竭评分最高(图 3C)。
功能比较显示,CD8+ T细胞亚群的细胞毒性从LME1到LME2再到LME3呈梯度下降,而LME2的细胞耗竭程度最高(图3D)。细胞因子和趋化因子程序(包括IFN-α和IFN-γ)的活性也反映了这些模式。综上所述,这些结果表明,LME亚型间T细胞的异质性既体现在组成上,也体现在功能上,反映了亚群比例以及细胞毒性和耗竭状态的差异。LME2中显著的耗竭特征进一步提示,针对逆转T细胞耗竭的免疫检查点阻断疗法可能更有效。
巨噬细胞进一步细分为 Mac_SPARC、Mac_STAB1、Mac_MARCO 和 Mac_CHIT1 四种亚型。Mac_SPARC 在 LME2 中的丰度高于 LME1,而 Mac_MARCO 在 LME1 中显著富集(图 3E)。值得注意的是,这四种亚型可根据两种经典标志物MRC1(编码 CD206)和FOLR2的表达进行区分,分别为 Mac_SPARC (FOLR2+MRC1−)、Mac_STAB1 (FOLR2+MRC1+)、Mac_MARCO (FOLR2−MRC+) 和 Mac_CHIT1 ( FOLR2−MRC1−)(图 3F-G)。他们还观察到巨噬细胞内FOLR2和TREM2的表达互斥,这与先前在实体瘤中的报道一致。
巨噬细胞亚型表现出显著的功能异质性。在典型的M1/M2极化模型中,CD206是M2表型的经典标志物。因此,CD206−Mac_SPARC和Mac_CHIT1表现出较高的M1评分,而CD206+Mac_MARCO则与较高的M2评分相关(图3H)。值得注意的是,Mac_MARCO在血管生成、吞噬作用和抗原呈递方面得分最高,表明其在淋巴微环境(LME)中发挥着核心枢纽作用(图3H)。虽然Mac_SPARC和Mac_MARCO均强烈影响辅助性T细胞(Th)的分化,但Mac_SPARC倾向于Th1,而Mac_MARCO倾向于Th2,这与其各自的M1/M2表型相符(图3H)。免疫应答富集分析 (IREA) 鉴定出这些巨噬细胞状态的不同上游细胞因子,其中 TNF-α 与 Mac_SPARC 相关,IL-4/IL-13 与 Mac_MARCO 相关。作为典型的 Th2 细胞因子,IL-4 和 IL-13 促进 CD206 表达并驱动替代性 (M2) 巨噬细胞活化。基因调控网络分析同样揭示了不同亚群中假定的上游调控程序存在异质性。
比较所有髓系亚型与LME亚型时,LME1表现出普遍较高的血管生成和中性粒细胞趋化性,这与其独特的细胞组成相符(图3J)。相比之下,T细胞和淋巴细胞的趋化性随主要巨噬细胞亚型而变化,例如LME1中Mac_MARCO的信号更强,而LME2中Mac_SPARC的信号更强,表明髓系亚型功能与LME组成之间存在相互依赖性(图3I)。
为了鉴定基质细胞的多样性,他们对成纤维细胞和内皮细胞进行了精细的亚群聚类。成纤维细胞包括 Fibro_PI16、Fibro_CCL19 和 Fibro_LRRC15,其中 Fibro_CCL19 在 LME2 中的富集程度显著高于 LME1(图 3J)。结合先前的报道和 CytoTRACE2 推断的发育潜能,Fibro_PI16 可能代表一种能够分化为其他两个亚群的前体成纤维细胞状态。在功能上,Fibro_CCL19 显示出更高的抗原加工和呈递水平、淋巴细胞趋化性以及通过 NF-κB 介导的 TNF-α 信号传导,而 Fibro_LRRC15 则富集于 ECM 相关程序(图 3K)。利用亚群特异性基因特征,NicheNet 揭示了包括 TNF-NFκB2 在内的相互作用是 Fibro_CCL19 的上游驱动因子,并鉴定出 TGFB1/TGFB2 是 Fibro_LRRC15 的关键配体(图 3L-M)。PROGENy 推断的通路活性独立地支持了配体-受体程序的亚群特异性(图 3N)。基因调控网络分析表明,Fibro_CCL19 的特征是炎症相关转录因子,包括 IRF1、STAT1、NFKB1 和 NFKB2(图 3O)。结合这些结果和先前的报道,他们将 Fibro_CCL19 归类为炎症性成纤维细胞状态,将 Fibro_LRRC15 归类为肌成纤维细胞状态。
即使在同一成纤维细胞亚型中,其功能也因淋巴微环境(LME)亚型而异。Fibro_CCL19 和 Fibro_LRRC15 在 LME2 中通过 NF-κB 活性表现出更高的 TNF-α 信号传导,而在 LME1 中则表现出更高的细胞外基质(ECM)组织化程度,其他通路也呈现类似的模式(图 3P)。因此,MALT 淋巴瘤中成纤维细胞的异质性体现在亚群特性和 LME 背景两个方面。
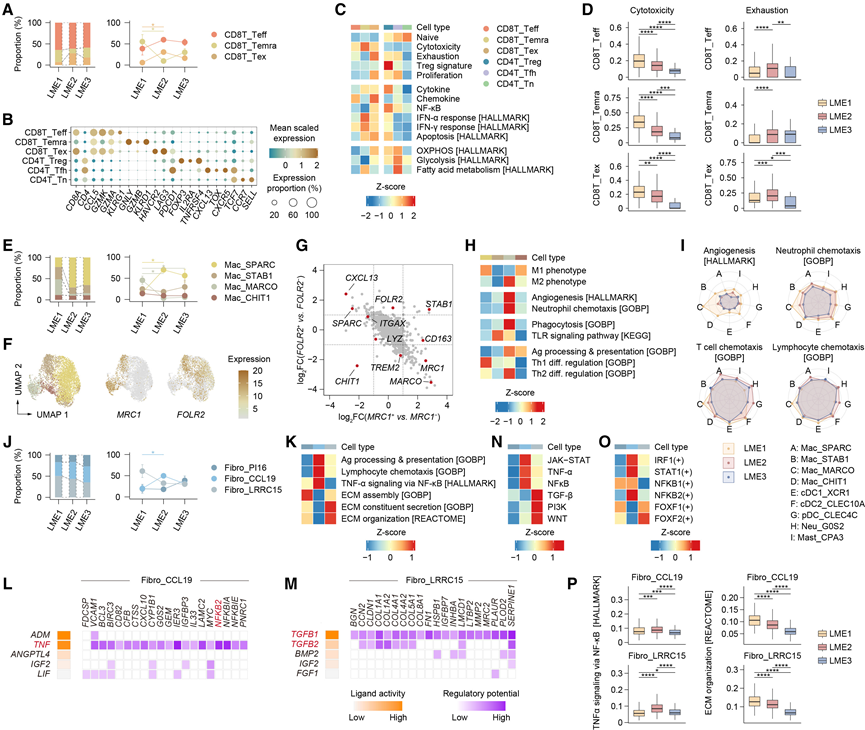
图3. LME亚型中免疫细胞和基质细胞的异质性。
(A) CD8+ T 细胞亚群在不同 LME 亚型中的相对比例(左图)。折线图显示各 LME 亚型中不同样本间亚群比例的差异(右图)。(B) 气泡图显示 T 细胞亚群中标记基因的表达。(C) 热图显示了T细胞亚群中代表性功能特征的富集情况。(D) 箱线图显示了各 CD8+ T 细胞亚群在不同 LME 亚型中的特征评分。(E) 与(A)相同的可视化,但针对巨噬细胞。(F) 通过 snRNA-seq 分析的5213 个巨噬细胞的 UMAP 图,以及MRC1和FOLR2的表达模式。(G) 与MRC1和FOLR2表达相关的代表性基因表达谱。(H) 热图显示巨噬细胞亚群中代表性功能特征的富集情况。(I) 雷达图显示了不同髓系细胞亚群在不同 LME 亚型中的代表性通路的变化。(J) 与(A)相同的可视化,但针对成纤维细胞。(K) 热图显示了成纤维细胞亚群中代表性通路的富集情况。(L) 热图显示了排名靠前的配体对 Fibro_CCL19 特征基因的调控潜力。(M) 与(L)相同的可视化,但针对的是 Fibro_LRRC15。 (N) 热图显示了成纤维细胞亚群中代表性的PROGENy衍生通路活性。(O) 热图显示了成纤维细胞亚群中代表性的pySCENIC衍生转录因子活性。(P) 箱线图显示了 Fibro_CCL19 和 Fibro_LRRC15 在不同 LME 亚型中的特征评分。
04
不同LME亚型中独特的细胞模块
在确定了不同LME亚型中的亚群异质性后,他们整合了所有亚群,并利用它们相对丰度的相关性,在MALT淋巴瘤中鉴定了细胞模块(CM)。CM1包含CD8T_Tex、CD4T_Treg、CD4T_Tfh、DC、Mac_SPARC、Fibro_CCL19、Endo_lymphatic和FDC_CR2,而CM2包含血管内皮细胞亚群、Neu_G0S2、Mac_MARCO和Peri_PDGFRB(图4A)。CM1在LME2中更为显著,而CM2在LME1中更为显著,这与这些LME亚型的特征一致(图4B)。
CellChat 分析显示,LME1 和 LME2 的整体细胞间相互作用强度强于 LME3,这可能反映了 LME3 细胞组成较为稀疏(图 4C)。在模块内部,CM1 成分之间的相互作用在 LME2 中最强,而 CM2 成分之间的相互作用在 LME1 中最强,这与模块比例相符(图 4C)。相对信息流表明,基质相关通路(包括 VEGF、TGF-β、PDGF 和胶原蛋白)在 LME1 中更为显著,而与淋巴细胞和 B 细胞活化相关的通路(包括 ICAM、CXCL、CD40 和 BAFF)在 LME2 中更为显著(图 4D)。在亚群水平上,Mac_SPARC、FDC_CR2 和 Fibro_CCL19 在 LME2 中的相互作用强于其他 LME 亚型,而 Neu_G0S2 和 Mac_MARCO 在 LME1 中的相互作用更强(图 4E)。对于恶性B细胞活化,关键通路CD40和BAFF信号均在LME2中向B_Mal细胞达到峰值。然而,这两个通路的来源不同。CD40信号主要来自其他细胞类型的旁分泌,而BAFF信号则主要来自B_Mal细胞的自分泌,表明二者的活化依赖性不同。
他们进一步研究了模块内不同LME亚型之间的配体-受体差异。在CM1中,CXCL、ICAM和CD40通路的总信号强度(由所有组成细胞类型的相对贡献之和计算得出)在LME2中更高(图4F)。在单个配体-受体对层面,连接CM1中包含的T细胞亚群的Mac_SPARC、FDC_CR2和Fibro_CCL19的相互作用在LME2中普遍更强(图4G)。在CM2中,VEGF和TGF-β的总信号强度在LME1中更高,其中Neu_G0S2的区分度最高(图4H)。 Neu_G0S2 和 Mac_MARCO 作为血管内皮细胞的信号发送者,尤其是 Neu_G0S2,在 LME1 中表现出广泛的信号增强,特别是通过 VEGFA 相关配体-受体对的信号传导,从而解释了 LME1 中观察到的血管生成能力增强(图4I)。鉴于 LME1 在胃 MALT 淋巴瘤中占主导地位,且HP感染在胃 MALT 淋巴瘤的发生中起着核心作用,他们进一步研究了中性粒细胞与 HP 感染相关基因特征以及血管生成特征之间的关系。本研究中胃 MALT 淋巴瘤样本的分析,结合两个独立的转录组微阵列数据集,一致揭示了中性粒细胞相关特征、HP 感染相关基因特征和血管生成相关通路之间存在正相关性。总而言之,细胞间相互作用分析表明,驱动淋巴细胞募集和功能激活的亚型特异性相互作用,以及血管生成,塑造了 LME 亚型之间独特的细胞景观。
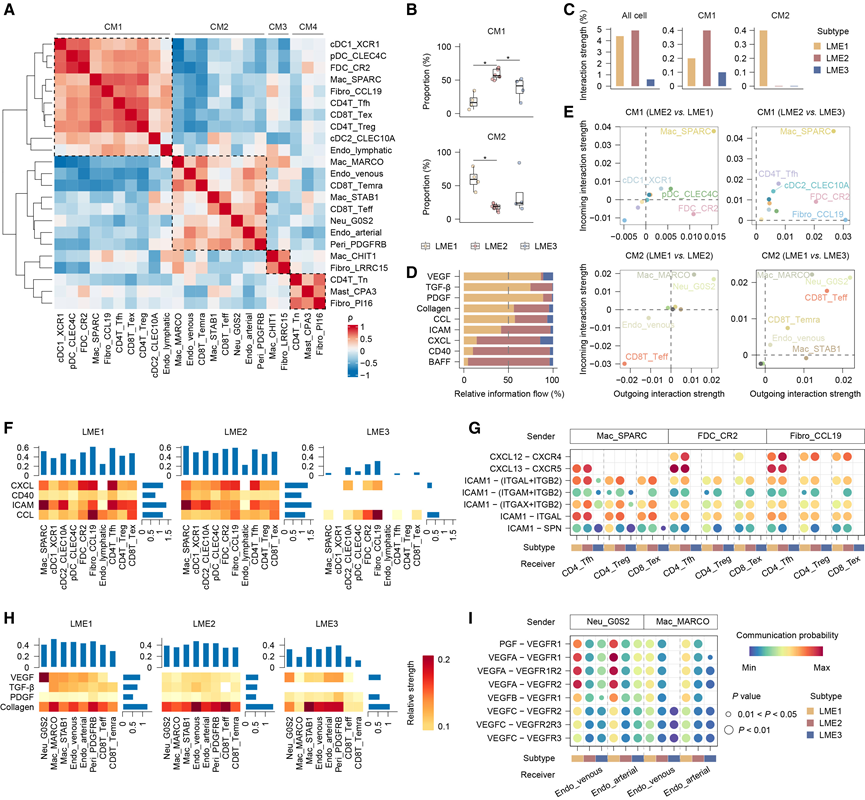
图4. 蜂窝模块和细胞间通信网络。
(A) 热图显示了MALT淋巴瘤样本中亚群比例的成对相关性。(B) 箱线图显示了各CM在LME亚型中的比例。(C) 柱状图比较了 LME 亚型之间的细胞间相互作用强度。(D) 代表性通路相对信息流突显了不同 LME 亚型中每条通路的重要性差异。(E) 散点图显示输出和输入交互强度,便于可视化 LME 亚类型之间的亚群体水平差异。(F) 热图(左下)显示不同LME亚型中CM1成分在CXCL、CD40、ICAM和CCL通路上的相对总体相互作用强度。柱状图(上)显示每种细胞类型在这些通路上的相对强度总和。柱状图(右)显示每种细胞类型中每种通路的相对强度总和。(G) 气泡图显示了 CM1 内不同细胞间相互作用中 CXCL 和 ICAM 通路中配体-受体对的通讯概率。(H) 与(F)相同的可视化,但针对 VEGF 通路中的 CM2 成分。(I) 气泡图显示了 VEGF 通路中配体-受体对在 CM2 内不同细胞间相互作用中 LME 亚型的通讯概率。
05
细胞间相互作用的空间映射
为了解析空间组织结构,他们对九个FFPE样本(每个LME亚型三个)进行了10倍Visium空间转录组学分析。这些样本的选择旨在确保LME之间的均衡代表性,并包含来自多个结外位点的样本(图5A)。利用cell2location软件将snRNA-seq数据整合,实现了细胞类型分布的空间分辨映射。共定位分析鉴定了五个空间因子(SF),这些因子捕捉了亚群共现的重复模式。CM1相关的亚群主要富集于SF2和SF3,而CM2相关的亚群则映射到SF5,这支持了每个模块内的空间共定位(图5B)。MISTy衍生的相互作用群落显示出类似的组织结构。空间模式在不同的SF之间存在差异。SF4在基质区域富集,而SF2和SF5分别与淋巴细胞和内皮细胞微环境相关,在微环境中分布更为分散(图5C)。与模块比例一致,SF2和SF3在LME2中更为显著,而SF5在LME1中更为显著(图5D)。代表性切片显示,CM1细胞类型优先定位于B_Mal附近,而CM2细胞类型虽然在内部存在共定位,但总体上距离B_Mal更远(图5E-F)。
为了分析这些空间关系,以CD8T_Tex+斑点为例,计算了每个B_Mal +斑点到周围CD8T_Tex +斑点的最近距离。在LME2中,这些距离较短,并且B_Mal相对于CD4T_Tfh、FDC_CR2和Mac_SPARC也观察到类似的接近程度(图5G)。与其他髓系细胞亚群相比,Neu_G0S2和Mac_MARCO的位置更靠近血管内皮细胞,这与其在血管生成中的作用相符。
他们进一步评估了配体表达与细胞间距离的关系。以 CD4_Tfh 为例,他们分析了每个斑点到 CD4T_Tfh+斑点的最近距离,并将其与斑点内的配体表达进行相关性分析。在 LME2 中,CD40LG与 CD4_Tfh、Mac_SPARC 和 FDC_CR2 的距离相关性较弱,表明这些细胞亚群附近区域的配体表达较高。类似地,在 LME1 中,VEGFA与 Neu_G0S2、Mac_MARCO、Endo_venous 和 Endo_arterial 的距离相关性也较弱(图 5H)。MISTy 分析进一步支持了这些空间关联,该分析发现 LME2 中 CD4T_Tfh 和CD40LG之间存在显著的斑点内相关性,并在 LME1 中突显了斑点内及其邻近区域的血管生成信号。
为了验证不同LME亚型中细胞群的空间关系,他们对15个样本进行了多重免疫荧光(mIF)染色,每个亚型选取5个代表性病例。鉴于MALT淋巴瘤LME中恶性B细胞的数量优势,他们将PAX5 +细胞归类为恶性B细胞(图5 I)。在LME2中,恶性B细胞倾向于更靠近FOXP3+ CD4+ T细胞和PD-1+ CD8 + T细胞(图5 J)。基于snRNA-seq衍生的标记定义和既往报道,他们将CD11b+ CD15+细胞注释为Neu_G0S2,将CD206+ MARCO+细胞注释为Mac_MARCO(图5 K)。定量分析显示,CD31 +血管内皮细胞在LME1中最为丰富,其中Neu_G0S2和Mac_MARCO均更靠近内皮细胞,这与空间转录组学结果一致(图5L)。此外,VEGFA+Neu_G0S2和Mac_MARCO在LME1中的频率更高,并且这些细胞群中的VEGFA +亚群比VEGFA-亚群更靠近内皮细胞,这与VEGFA的促血管生成作用相符。
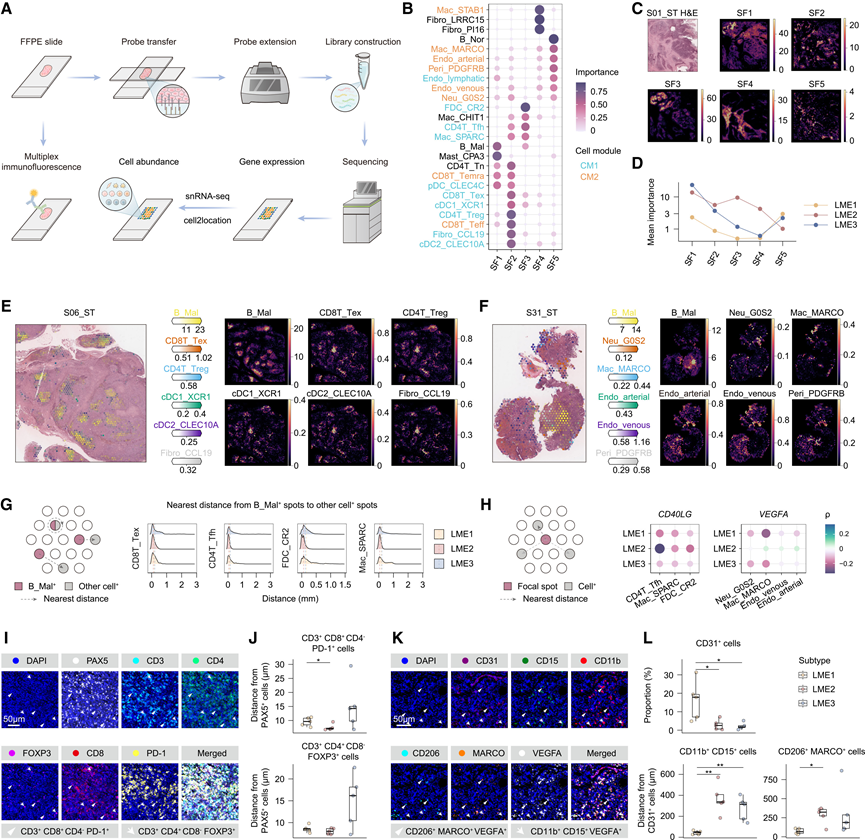
图5. 细胞间相互作用的空间组织。
(A) 基于 FFPE 切片的空间 RNA-seq 和 mIF 的示意图。(B) 气泡图显示每个SF中细胞亚群的重要性。(C) 代表性切片中 H&E 染色和 SF 分布的空间可视化。(D) 折线图显示三个 LME 亚型中每个 SF 的平均重要性。(E) 利用 cell2location 方法将 CM1 成分和 B_Mal 中的细胞类型空间映射到空间 RNA-seq 玻片上。估计的细胞丰度用颜色强度表示,所有细胞类型(左)以及每种细胞类型(右)。(F) 与(E)相同的可视化,但针对 CM2 成分和 B_Mal。(G) 示意图(左)展示了B_Mal +斑点(红色)与其他细胞类型阳性斑点(灰色)之间最近距离的计算方法。如果两个斑点重叠,则最近距离定义为0。脊线图(右)描绘了B_Mal +斑点与指定细胞类型之间最近距离在不同LME亚型中的分布。(H) 示意图(左)展示了焦点(红色)与细胞类型阳性点(灰色)之间最近距离的计算。(I) mIF染色代表性图像显示了LME中的T细胞亚群。(J) 箱线图显示每个LME亚型样本中PAX5+细胞到CD3+ CD8+CD4− PD-1+细胞(上图)和CD3+ CD4+ CD8− FOXP3+细胞(下图)的平均距离。(K) mIF染色代表性图像显示了LME中的血管内皮细胞和相关的髓系亚群。(L) 箱线图显示了每个LME亚型样本中CD31+细胞的比例(上图)以及CD31+细胞与CD11b+ CD15+细胞(左下图)和CD206+ MARCO +细胞(右下图)的平均距离。
+ + + + + + + + + + +
结 论
本研究报告了来自多个解剖部位的89例MALT淋巴瘤样本,基于多组学数据获得的细胞类型特异性特征,对来自本研究队列和两个独立队列的120个整体转录组进行整合分析,鉴定出三种LME亚型:间质型、炎症型和耗竭型。单细胞和空间映射进一步揭示了这些亚型之间不同的细胞组成和相互作用网络,尤其是在B细胞活化和血管生成方面,提示了不同的生物学状态和亚型特异性的治疗弱点。本研究建立了一个基于LME的MALT淋巴瘤分类框架,具有生物学和临床意义。
+ + + + +



