

 English
English文献解读|Signal Transduct Target Ther(52.7):单细胞鉴定和验证非小细胞肺癌 中人循环 Treg 亚型/状态 Treg fci
✦ +
+
论文ID
原名:Single-cell identifies and validates human circulating Treg subtype/state Tregfci in non-small cell lung cancer
译名:单细胞鉴定和验证非小细胞肺癌 中人循环 Treg 亚型/状态 Treg fci
期刊:Signal Transduction and Targeted Therapy
影响因子:52.7
发表时间:2026.06.03
DOI号:10.1038/s41392-026-02677-6
背 景
循环T细胞对于维持全身免疫稳态、协调器官间免疫反应以及调节局部微环境至关重要。随着单细胞转录组分析(scRNA-seq)和细胞身份标记基因panel(ciMGP)的出现,T细胞亚型/状态(Ts/s)和身份的发现迅速扩展。空间转录组学分析(ST)表明,肿瘤微环境(TME)内Ts/s的分布和动态变化与癌症恶性程度、肿瘤侵袭、抗肿瘤免疫微环境的形成以及治疗反应密切相关。循环T细胞在稳定全身免疫功能、模拟空间化的微环境防御以及反映器官间通讯方面发挥着关键作用。scRNA-seq能够精细分析活化或过度增殖的T细胞及其受体的丰度、多样性和特征,从而为接受免疫检查点阻断剂治疗的黑色素瘤患者提供免疫相关不良事件严重程度和进展的生物标志物。scRNA-seq定义的Ts/s为T细胞图谱、疾病特异性T细胞模式以及Ts/s之间的相互作用提供了新的见解,有望转化为血液学临床生物化学领域的临床应用。然而,由于成本负担、技术复杂性、法规、设备认证、标准化、可重复性、数据解读、生物信息学安全、医生专业知识差异以及法律框架等问题,scRNA-seq定义的Ts/s的临床转化仍然面临挑战。其中,主要障碍在于精确选择T细胞,而T细胞发育、定位、调控、功能可塑性和代谢异质性的多方面特性使得这一过程变得复杂。
实验设计
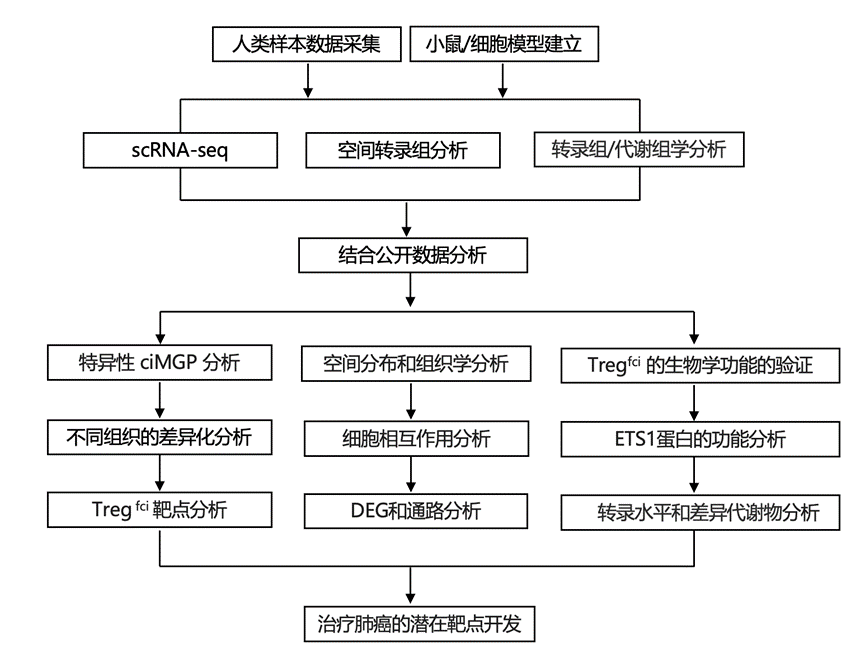
结 果
01

基于 rOER 的 Ts/s ciMGP 的选择
为了启动T细胞ciMGP的筛选,研究团队从自身数据、11个自建的scRNA-seq数据库以及公开的scRNA-seq数据库中收集了Ts/s ciMGP(图1a)。经过初步筛选,我们筛选出约300个候选ciMGP,用于后续的scRNA-seq分析中的T细胞注释。去除物种和组织间的重复条目或同义基因集后,他们保留了170-262个ciMGP用于后续分析。为了确保命名统一,他们根据标准化的标记或特征性功能基因对相应的T细胞亚群进行了统一的重命名,通过评估阈值、重叠数量、区域重叠表达率(rOER)分析和ciMGP分选对人血样本和组织中的T细胞ciMGP进行了特异性评估(图1a)。
为了确定每个ciMGP在scRNA-seq数据注释中的特异性,他们对目标ciMGP mRNA表达的重叠区域进行了量化,并使用箱线图进行了可视化(图1b),将对应Q3和Q1之间的目标面板箱定义为非目标ciMGP的可重复范围。阈值设定为每个样本通过scRNA-seq鉴定的ciMGP总数的10%,并根据检测到的亚型数量进行调整。根据重复性范围内重叠的 ciMGP 比例,将ss-ciMGP定义为数量>30% < 阈值的 ciMGP(图1c),将 sa-ciMGP 定义为数量>30% > 阈值且>60% < 阈值的 ciMGP(图1d),将 sr-ciMGP 定义为数量>60% > 阈值的 ciMGP(图1e)。
为了阐明特定ciMGP在正常PBMC中的存在情况,他们从一组健康个体中分离出89982个人外周血单核细胞 (PBMC)进行聚类分析,其中检测到123个Ts/s。在已鉴定的ciMGP中,29个(39%)定义为ss-ciMGP,8个(11%)定义为sa-ciMGP,37个(50%)定义为sr-ciMGP。
为了鉴定非小细胞肺癌(NSCLC)特异性T细胞ciMGP,本研究招募了初次就诊时年龄在47至78岁之间的患者,疾病分期为I至III期。对NSCLC患者术前和术后外周血样本以及肿瘤和邻近肺组织样本中的122747个T细胞进行了scRNA-seq(图1f)。分别分析了术前和术后血液样本中的约9915个和11675个T细胞,以及分别分析了NSCLC组织和邻近肺组织样本中的11742个和10268个T细胞(图1g)。初筛后T细胞ciMGP的检出率得到验证,术前或术后血样中分别检测到93个或99个T细胞ciMGP,而在NSCLC或para-NSCLC组织中分别检测到117个或104个T细胞ciMGP(图1h)。其中,80个ciMGP出现在所有样本中,13个出现在术后血样、NSCLC和非小细胞肺癌癌旁组织 (para-NSCLC)样本中,4个出现在术前和术后血样以及NSCLC样本中,如图1h所示。ciMGP的数量和 rOER 值分为 ss-ciMGP、sa-ciMGP和 sr-ciMGP。术前或术后血液中 ss-/sa-/sr-ciMGP 的数量分别为 63/9/21 或 65/15/19,在NSCLC组织中分别为 78/19/20,在para-NSCLC组织中分别为 58/21/25。
为了评估T细胞ciMGP的疾病特异性,他们比较了NSCLC患者和健康个体在不同解剖部位和时间点的ciMGP的存在情况和频率。与健康个体的ss-ciMGP相比,术前血液、术后血液、NSCLC组织和para-NSCLC组织中分别缺失约41%、38%、21%和45%的Ts/s。在所有样本类型中,ss-ciMGP的数量均显著增加,分别达到健康对照组基线值的159%、162%、190%和145%(图1i)。
在Ts/s中,有26个与NSCLC患者的无进展生存期(PFS)、总生存期(OS)和术后无进展生存期(PPS)显著相关。肺癌患者的OS在Ts/s水平高低之间存在显著差异。其中,NSCLC样本和术前血液中循环T细胞亚型Tregfci的数量和表达显著升高,而术后血液中则未检测到(图1j)。与低表达的NSCLC患者相比,5g-Tregfci或3g-Tregfci高表达的NSCLC患者的OS显著更差(图1k),这在肺腺癌(LUAD)患者中也观察到。为了评估表达CD4、CD45、FOXP3、CTLA4和IL2RA的Treg(5g-Tregfci)或表达FOXP3、CTLA4和IL2RA 的Tregfci对多发性癌症患者长期生存率的潜在影响,他们重新评估发现,5g- Tregfci或 3g-Tregfci ciMGP高表达的患者生存率较差,且 5g-Tregfci和 3g-Treg fci之间无显著差异。Tregfci之所以选为 NSCLC 特异性 Ts/s 之一,是基于其 ciMGP 的高度特异性、独特的表达模式(其 ciMGP基因表达量是对照组的三倍以上)以及与生存率的强相关性。
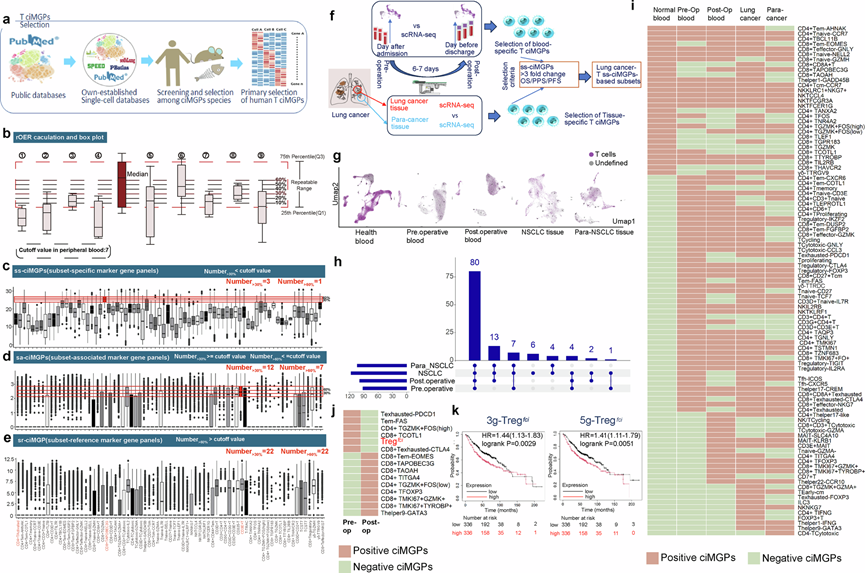
图1. T细胞特异性标记基因(ciMGP)在不同免疫区室和临床条件下的工作流程和分类。
(a) 共300个候选标记基因panel,这些panel与来自不同来源和筛选物种(人、小鼠和斑马鱼)的不同T细胞亚群和功能状态相关。(b) 使用区域重叠表达率(rOER)评估ciMGP特异性的策略。(c) 亚群特异性ciMGP(ss-ciMGP)是指在少于阈值亚群中出现且rOER>30%的ciMGP。(d) 亚群相关 ciMGP(sa-ciMGP)是指在超过阈值数量的亚群中表达 rOER 值介于 30% 至 60% 之间的ciMGP。(e) 亚群参考 ciMGP(sr-ciMGP)是指在超过阈值数量的亚群中表达且 rOER > 60% 的 ciMGP,代表广泛表达但鉴别能力有限的 ciMGP。(f) 循环 T ciMGP 验证研究设计的示意图。(g) 基于 rOER 定义的亚群特异性的 T 细胞亚群分布的 UMAP 可视化图。(h) Upset图展示了在非小细胞肺癌 (NSCLC) 患者血液和肺组织样本中鉴定的 T ciMGP 的交集。(i) 热图显示了术前或术后血液、NSCLC 或para- NSCLC 肺组织中是否存在 ss-ciMGP。(j) 热图显示术前或术后血液中 rOER 定义的 T 细胞 ciMGP 的出现和变化。(k)生存分析。
02
所选靶点 Tregfci的验证
为了进一步验证人 Tregfci的特异性,他们招募了 45 名年龄在 20 至 50 岁之间的健康志愿者,对其PBMC进行scRNA-seq,并与公共数据集中的结果进行比较。在鉴定的 Ts/s 中,有 123 个 (63.1%) 在本研究队列 PBMC 中检测到,其中 72 个 Ts/s 是本研究队列特有的(图2a)。本研究队列中约 71% 的 Ts/s 为单链 ciMGP,且其大小高于公共数据库中的比例(图2b),而本研究队列和公共数据库中 Ts/s 的功能组成模式相似(图2c)。在这些 Ts/s 中,NK 细胞和 NKT 细胞占前 20 个变化最显著的 Ts/s 的 45%。 UMAP可视化结果显示,血液中T细胞组成与肿瘤负荷状态密切相关(图2d),在NSCLC组织、邻近非肿瘤组织、术前术后血液以及健康受试者的血液中,T细胞亚型分布存在显著重叠和独特性(图2e)。3g-Treg fci是少数在肿瘤切除前后循环中出现或消失的亚型之一(图2f),并且在NSCLC组织中显著富集(图2g)。比较不同Treg亚群发现,Tregfci中ciMGP的表达量是其他亚群的三倍以上,且NSCLC组织和邻近非小细胞肺癌(para-NSCLC)组织中5g-Tregfci等亚群的丰度存在明显差异。通过分别评估5g-Tregfci和3g-Tregfci的出现和定位,比较了二者空间分布的差异和一致性。来自公共数据集的年龄 <50 或 ≥50 的组之间5g-Tregfci或 3g-Tregfci的出现频率相似。
为了验证血液中 Tregfci的可检测性,他们对健康志愿者、健康志愿者的 5g-Tregfci样本、NSCLC患者的 3g-Tregfci样本以及NSCLC患者的 5g-Treg fci样本进行了流式细胞术染色,检测经典 Treg(CD45+CD3+CD4+CD25+CD127−)和 Tregfci标记物。结果显示,与健康对照组相比,NSCLC 患者血液中的Tregfci细胞群显著增加(图2h)。为了评估 Tregfci 的疾病和器官/组织特异性,他们选取了 64 种 T 细胞(包括 Tregfci)在组织/器官以及肺部或肺外疾病中进行了评估。为了确定NSCLC的特异性,他们比较了不同肺部疾病中Tregfci的数量,发现NSCLC组织中Treg fci/总T细胞的比值显著高于para-NSCLC组织、肺腺癌(LUAD)、、慢性阻塞性肺疾病(COPD)、特发性肺纤维化(IPF)和系统性硬化症(SSC)(图2i)。为了评估年龄对Treg fci丰度的影响,他们利用流式细胞技术从另外76例NSCLC患者的外周血中分离并检测了5g-Tregfci的数量,其中包括43例年轻患者和33例老年患者。结果发现,年轻和老年NSCLC患者的5g-Tregfci数量均显著高于年轻和老年健康对照组(图2j)。年轻和老年健康对照组之间无显著差异。他们对年轻和老年NSCLC及para-NSCLC组织中的Tregfci标志物及其靶蛋白进行了免疫组织化学染色分析。与年轻患者相比,老年患者肿瘤内及肿瘤周围组织中Treg细胞和Treg聚集体的评分以及ETS原癌基因1(ETS1)蛋白的表达强度均较高。他们将NSCLC中T细胞亚群的ciMGP模式与其他肺部疾病进行了比较(图2k)。在17个器官/组织中,Tregfci主要见于乳腺、宫颈、皮肤、小肠和口腔组织。在分析的31种肺外疾病中,Tregfci也见于其他肺部疾病。
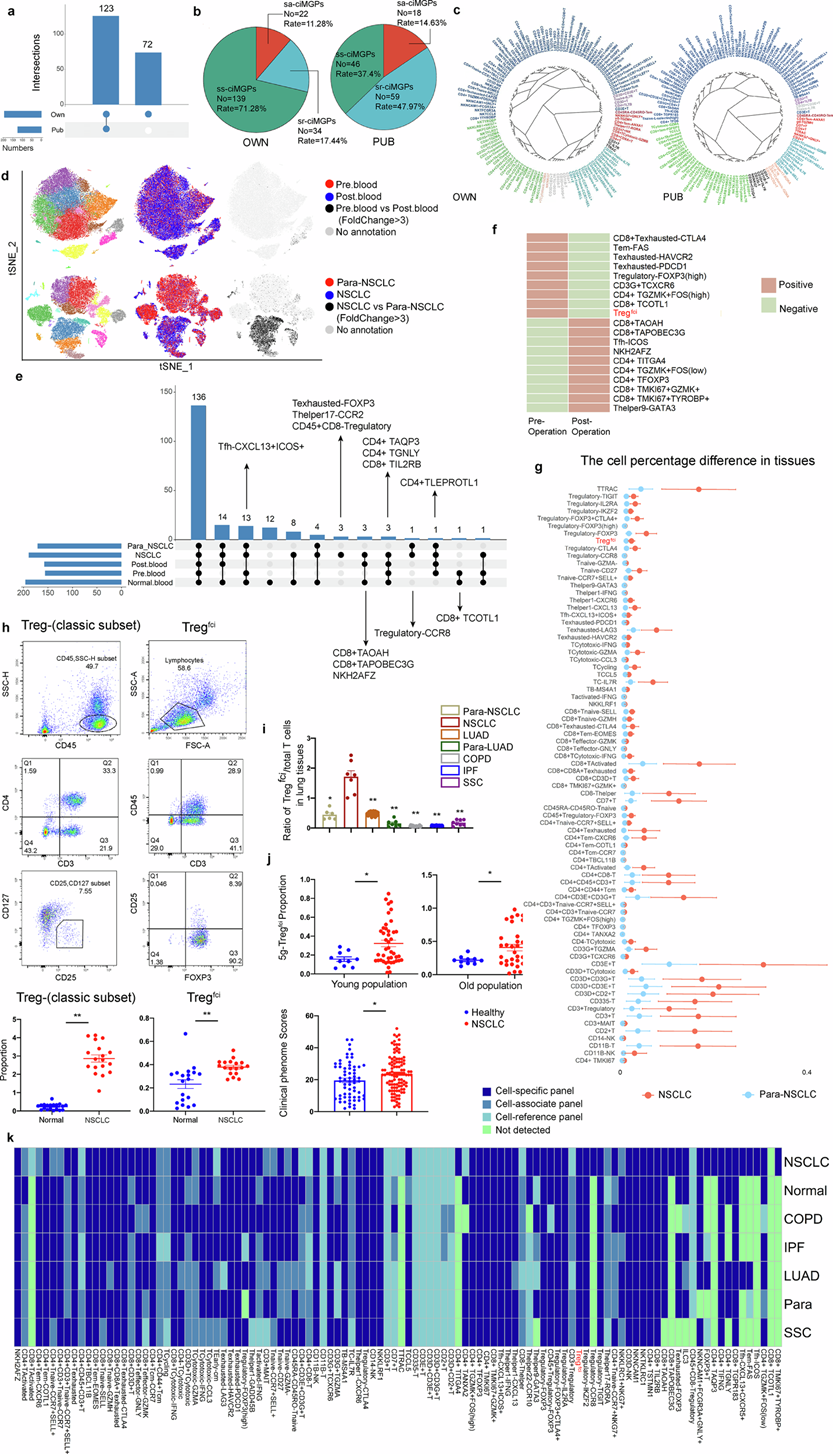
图2. 对健康个体、NSCLC患者、肺部疾病和肺外疾病中的 T 细胞 ciMGP 进行分析。
(a) 在 PBMC 中鉴定的 T 细胞 ciMGP 的交集图。(b) 为使用 rOER 工具确定的在 PBMC 中鉴定的 T 细胞 ciMGP 的分类饼图。(c) 使用 rOER 工具可视化 195 和 123 个 T 细胞 ciMGP 聚类的系统发育树。(d) 显示来自 NSCLC 患者血液和肺组织的整合 scRNA-seq 数据。(e) 展示了 NSCLC 患者和健康志愿者血液和肺组织样本中已识别的 T ciMGP 的交集。(f) 热图显示了 NSCLC 患者术前或术后血液样本中 T ciMGP 的存在(橙色)或缺失(绿色)。(g) 点图描绘了 NSCLC 组织之间变化超过 3 倍的 T ciMGP 的比例。(h-j) 流式细胞分析。(k) 热图比较了 108 个选定的 T ciMGP 在 NSCLC 组织与正常肺、LUAD、para- LUAD、COPD、IPF 和 SSC 中的分布和变异性。
03
空间和功能分析
为了阐明 Tregfci的空间分布和生物学特征,我们对取自健康供体和NSCLC患者的配对肺组织切片进行了ST分析(图3a)和 HE 染色(图3b)。Tregfci、ILC3和 CD8 T细胞毒性-IFNG弥漫分布于肺癌和癌旁组织中。Tregfci在 NSCLC 组织中显著富集,并且在肿瘤核心和邻近区域之间表现出明显不同的空间分布模式(图3a)。他们观察到 NSCLC 肿瘤中心区和外周区之间存在显著差异(图3c)。与其他癌症相比,非小细胞肺癌中心Tregfci ciMGP的表达更高,这证明了Tregfci的空间分布和外观的特异性。
肿瘤中心区域的Tregfci丰度显著低于周边区域(图3d)。Tregfci ciMGP在癌旁区富集,但在恶性区域显著下降,提示晚期肿瘤可能存在免疫逃逸机制。为了探究 Tregfci在不同癌症类型中的独特模式,他们评估了 Treg fci在 BRCA、结直肠癌(CRC)、肝癌(HCC)、卵巢癌(OV)和胰腺导管腺癌(PDAC)中的定位。在 6 例 BRCA 患者中的 2 例中,肿瘤与癌旁交界处的Tregfci密度显著高于肿瘤组织本身,甚至高于癌旁区域(图3e)。在 CRC(图3f)和 OV(图3g)中也观察到了类似的分布模式。相比之下,HCC(图3h)和PDAC(图3i)的界面区域的Tregfci密度显著高于癌旁组织。NSCLC和癌旁组织中Tregfci的 转录组比较显示,多种信号通路上调,包括原发性免疫缺陷、亚油酸代谢、抗原加工和呈递以及聚糖生物合成-硫酸乙酰肝素(图3j)。为了表征术前血液特异性T细胞/S细胞中Tregfci介导的细胞间通讯,他们评估了NSCLC和癌旁组织中存在的T细胞/S细胞的ciMGP。Tregfci与Tem-FAS、NSCLC中Treg fci自身以及Treg-FOXP3(high)细胞的相互作用最弱(图3k)。术前血液特异性T细胞亚群与在非小细胞肺癌旁组织(para-NSCLC)区域检测到的癌症ciMGP的相互作用显著上调,特别是癌症CXCL14与癌症干细胞之间的相互作用,相比于与其他细胞亚群的相互作用(图3l)。
在para-NSCLC组织中,Tregfci介导的与Tem-FAS以及Tregfci自身的相互作用较弱(图3l)。Tregfci ciMGP的表达也通过近期发表的中国免疫多组学图谱(CIMA)进行了验证。CIMA包含来自428名成年人的性别、年龄和遗传相关的免疫谱,并比较了超过1000万个免疫细胞。根据CIMA注释,IL2RA在ILC2亚群中高表达(图3m),CTLA4在MKI67+CD8+T细胞亚群中高表达(图3n),IFI44L在CD4 + T细胞亚群中高表达。Tregfci ciMGP在两个注释的Treg亚群中显著表达:CD4-Treg-FCRL3和CD4-Treg-FOXP3(图3o)。转换前记忆B细胞亚群JAM和DC_CLEC9A分别高表达IL2RA和CTLA4。为了精确定位Tregfci的空间分布,他们使用针对单个ciMGP蛋白的特异性抗体以及包含所有三种Tregfci标记物的多重抗体进行免疫染色。FOXP3阳性细胞主要定位于TME内(图3p),证实了Tregfci在肿瘤组织中的空间富集。多重染色显示,FOXP3⁺CTLA4⁺IL2RA⁺阳性细胞主要富集于肿瘤组织及其邻近区域(图3q)。
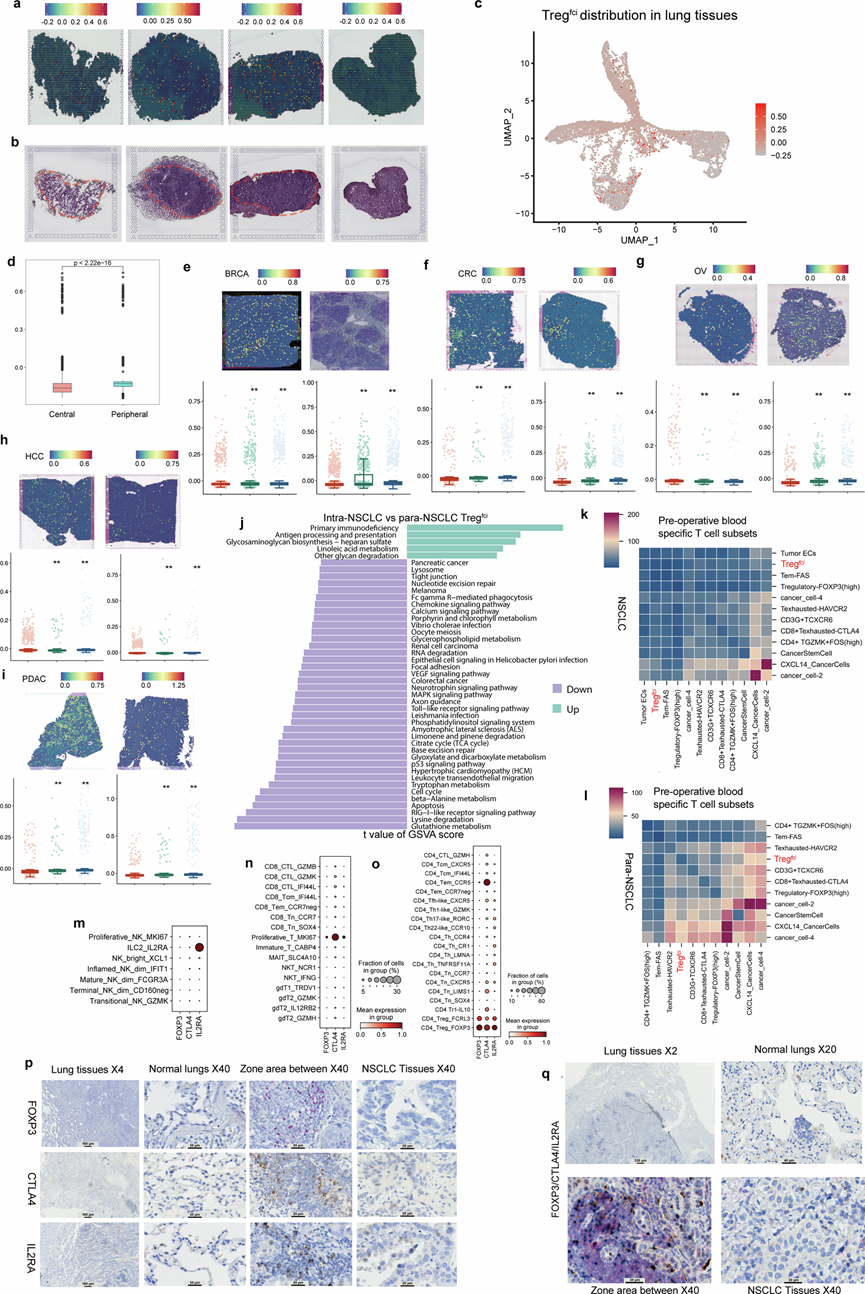
图3. 对非小细胞肺癌 (NSCLC) 和其他实体瘤中的Tregfci亚群进行空间和功能分析。
(a) 3 例 NSCLC 患者组织切片中Tregfci基因特征的基因集变异分析 (GSVA) 评分的空间转录组可视化。(b) NSCLC 和非 NSCLC 区域的代表性苏木精-伊红 (H&E) 染色图像和形态结构,以及 Tregfci的分布。(c) NSCLC 组织中转录组斑点的空间分布的 UMAP 可视化。(d) 箱线图比较NSCLC 组织中心区域和外周边缘区域Treg fci相关基因特征评分。(e-i) Tregfci特征评分。 (j) 基于 GSVA 的通路富集直方图,分析了非小细胞肺癌 (NSCLC) 和para- NSCLC 组织中 Treg fci亚群的差异表达基因 (DEG)。通过比较肿瘤和非肿瘤组织,鉴定出 44 个富集通路,用于表征 Treg fci亚群的功能差异。(k-l) CellPhoneDB 配体-受体相互作用分析。(m-o) 气泡图展示了来自 428 名健康个体(CIMA 队列)的 43 个已注释的 T 淋巴细胞亚群中 Tregfci ciMGP 的 mRNA 表达情况。(p) 免疫组化染色。(q) 多重免疫组化共表达染色。
04
空间富集分析
为了明确 Tregfci的功能和调控异质性,他们比较了Treg fci与术前和术后 T 细胞的转录组谱。通过与术前血液中其他 T 细胞进行比较,他们鉴定了Tregfci中上调和下调最显著的 10 个基因(图4b),以及术前血液中Tregfci与术后血液中总T细胞之间的差异(图4a)。在差异表达基因(DEG)中,与术前或术后 T 细胞相比,ETS1在术前 Treg fci中显著上调。ETS1、RGS1和TRBC2在术前血液的Tregfci中持续升高。他们通过比较定量频率(图4c)和分别以热图形式可视化 20 种术前和术后血液特异性 T 细胞亚群的差异表达基因谱(图4d),研究了术前血液特异性 T 细胞亚群间的差异。ETS1是 Tregfci中主要上调的基因。通过比较 NSCLC 的中心和外周肿瘤区域,测量了空间转录组样本所有点(图4e)和 Tregfci内(图4f)的 ETS1 表达值。
为了解ETS1下调的Treg fci在人体系统中的作用,他们构建了Treg ETS1-KD或Treg ETS1-NC的特殊人源化动物模型,发现移植PBMC两周后,人源化小鼠体内人CD45阳性细胞的比例超过25%,并对这些小鼠进行了肿瘤细胞接种。从第3天到第15天,他们观察到活细胞中人CD45阳性细胞的比例增加,而小鼠CD45阳性细胞的比例减少。为了更好地反映人体免疫生物学特性并避免同种异体反应的潜在影响,他们利用来自患者的PBMC和肺癌细胞构建了自体人源化类器官模型,并以1:2的比例每两周传代一次扩增类器官。类器官内的细胞形态似乎起源于肺癌细胞。
与肿瘤中心区域相比,ETS1在肿瘤外周区域的表达显著升高。空间分析显示,在6种肺外肿瘤中,ETS1主要富集于NSCLC与邻近癌旁组织的交界区域(图4g)。在其他癌症类型中,包括BRCA、CRC、HCC、OV和PDAC,肿瘤/癌旁交界区域和癌旁区域的ETS1表达均有所增加。在HCC组织中,与HCC和癌旁组织相比,ETS1在交界区域显著过表达。本研究阐明了在各种生理和病理条件下(包括健康供体、NSCLC和其他肺部疾病)16个已定义的Treg亚群中ETS1的表达情况。在健康外周血中,ETS1表达在 Treg CTLA4、CD45+ Treg FOXP3、Treg IKZF2或 Treg Tight中相对较高,但在CD3+ Treg 和 Treg FOXP3中较低(图4i)。
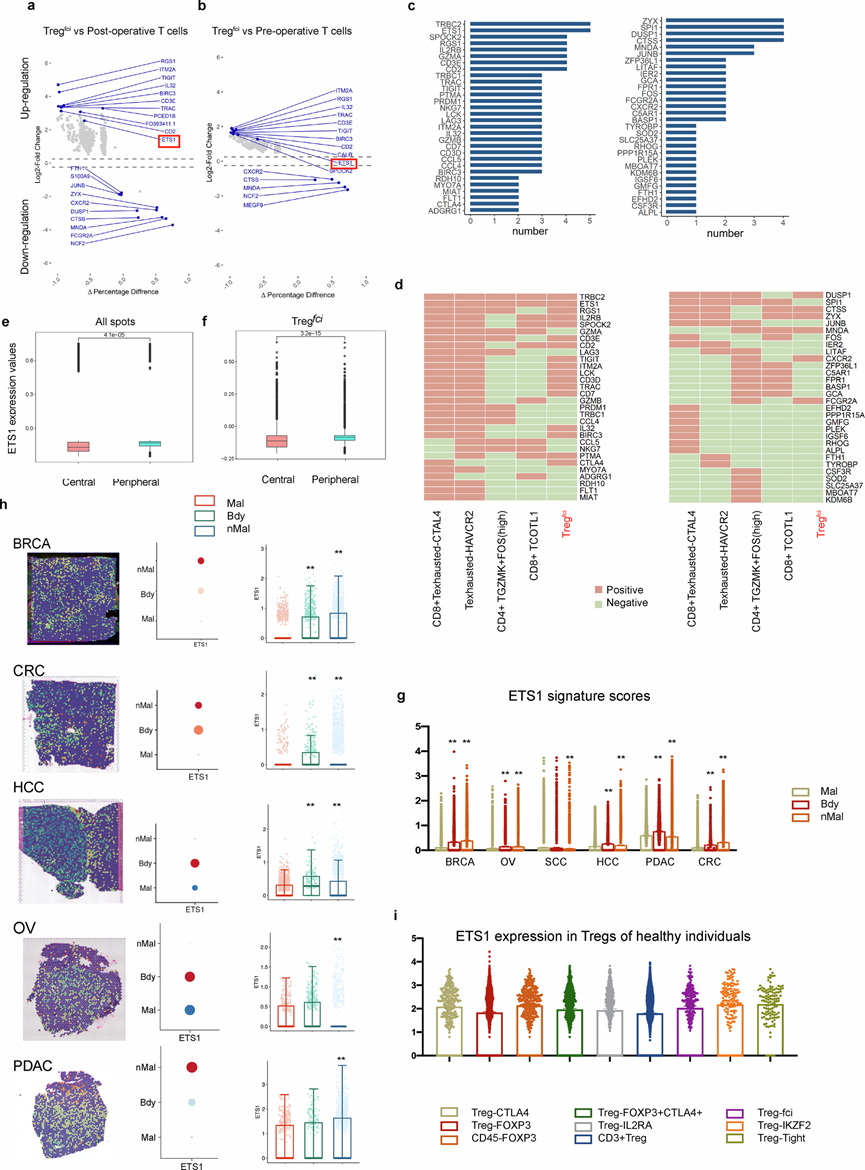
图4. ETS1相关的转录组图谱及Treg fci亚群在非小细胞肺癌(NSCLC)和多种免疫病理背景下的空间富集情况。
(a-b) 鉴定的DEG。(c) 柱状图展示了NSCLC患者术前血液特异性T细胞亚群与术后血液T细胞之间前30个DEG。(d) 热图展示了NSCLC患者术前血液特异性T细胞亚群与术后血液T细胞之间前30个上调/下调差异表达基因的表达谱。(e-f) 细胞的特征评分的空间分布。(g) 箱线图显示了通过SpatialTME分析的BRCA、CRC、HCC、OV、PDAC和SCC中ETS1在不同空间区域的表达情况。(h) 使用SpatialTME分析了BRCA、CRC、HCC、OV、PDAC和SCC中ETS1的空间分布。(i) 箱线图显示了ETS1的表达情况,以及在健康供体中16个Treg ciMGP中9个的鉴定结果。
05
Tregfci生物学功能的验证
为了评估每种 Tregfci ciMGP 在 T 细胞功能和相互依赖性中的作用,他们使用慢病毒 shRNA 载体选择性地下调了FOXP3、CTLA4和IL2RA的表达(分别记为 T FOXP3-KD、T CTLA4-KD和 T IL2RA-KD),或同时下调了 T 细胞中所有 ciMGP 的表达(Tfci-KD)。我们评估了细胞生物学功能和细胞间相互作用(图5a)。结果表明,与阴性对照 T NC相比,T FOXP3-KD、T CTLA4-KD和 Tfci-KD的细胞活力显著降低。经 PHA 处理的T CTLA4-KD(图5b)和 T IL2RA-KD在培养第 3 天即表现出增殖能力丧失。与T-NC对照组相比,在单独或联合敲低的细胞中,三个基因的表达水平显著降低,尤其是FOXP3(图5c)在T FOXP3-KD和Tfci-KD细胞中,以及IL2RA(图5d)和CTLA4(图5e)分别在T CTLA4-KD和Tfci-KD细胞中降低。与T NC对照组相比,PHA 预处理增加了所有T细胞群中IL2RA的表达。PHA处理增加了Tfci-KD细胞中FOXP3的表达,并降低了IL2RA和CTLA4的表达水平。
为了评估 Treg fci ciMGP在 T 细胞毒性作用中的调控作用,他们将 Tfci-NC或 Tfci-KD与 HBE 或 A549 细胞共培养,并分别进行或不进行 PHA 刺激。他们检测了不同浓度和时间点下 T 细胞的杀伤能力。使用结晶紫染色法观察残余靶细胞的活力和形态。他们选择 PHA 刺激后 A549 细胞与 T 细胞比例为 1:10 的培养条件(图5f),并评估下调 Tregfci ciMGP 对 T 细胞杀伤能力的影响。在这些敲低细胞中,Tfci-NC对 PHA 的敏感性最高,且与是否与 A549 细胞(图5f)或 HBE 细胞(图5g)共培养无关,因为无论是否用载体或 PHA 处理,Tfci-NC和 Tfci-KD的细胞活力均存在显著差异。当分别与浓度为1:1或1:10的HBE共培养时,Tfci细胞的存活率显著升高或降低(图5j)。Tfci-KD加剧了PHA处理的Tfci-NC细胞的毒性作用,这体现在与载体处理的Tfci-KD和Tfci-NC细胞的比较,以及载体处理和PHA处理的Tfci-KD和Tfci-NC细胞存活率下降程度的比较(图5g)。然而,敲低FOXP3或CTLA4并未导致载体或PHA处理下毒性的显著差异。此外,他们还检测了Tfci-NC对8种癌细胞系存活率的影响,发现与Tfci-NC联合载体或PHA处理相比,HBE细胞在载体刺激后与Tfci-KD共培养,以及A549、H460、H446和SPC-A1细胞在PHA刺激后与Tfci-KD共培养,其存活率均较低。LTEP细胞经载体和Tfci-KD处理后的存活率显著高于Tfci-NC联合载体或Tfci-KD联合PHA处理。而Tfci-KD联合载体或PHA处理的动态存活率则高于Tfci-NC。
细胞凋亡检测结果显示,与载体处理组相比, PHA刺激显著增加了Tfci-NC细胞的早期和晚期凋亡率。Tfci-KD细胞增加了基础凋亡率,并减弱了PHA诱导的Tfci-KD细胞的凋亡敏感性(图5k)。与A549或HBE细胞共培养可降低T细胞凋亡,尤其是晚期凋亡,与单独培养的T细胞相比,晚期凋亡率下降了近10倍。除PHA处理的Tfci-NC细胞外,与Tfci-NC或Tfci-KD细胞共培养的A549/HBE细胞中PHA诱导的凋亡极少。A549与PHA刺激的 Tfci-NC或 Tfci-KD共培养时,细胞凋亡率降低(图5l)。PHA处理的Tfci-KD增加了共培养的 HBE 细胞的早期凋亡率(图5m)。
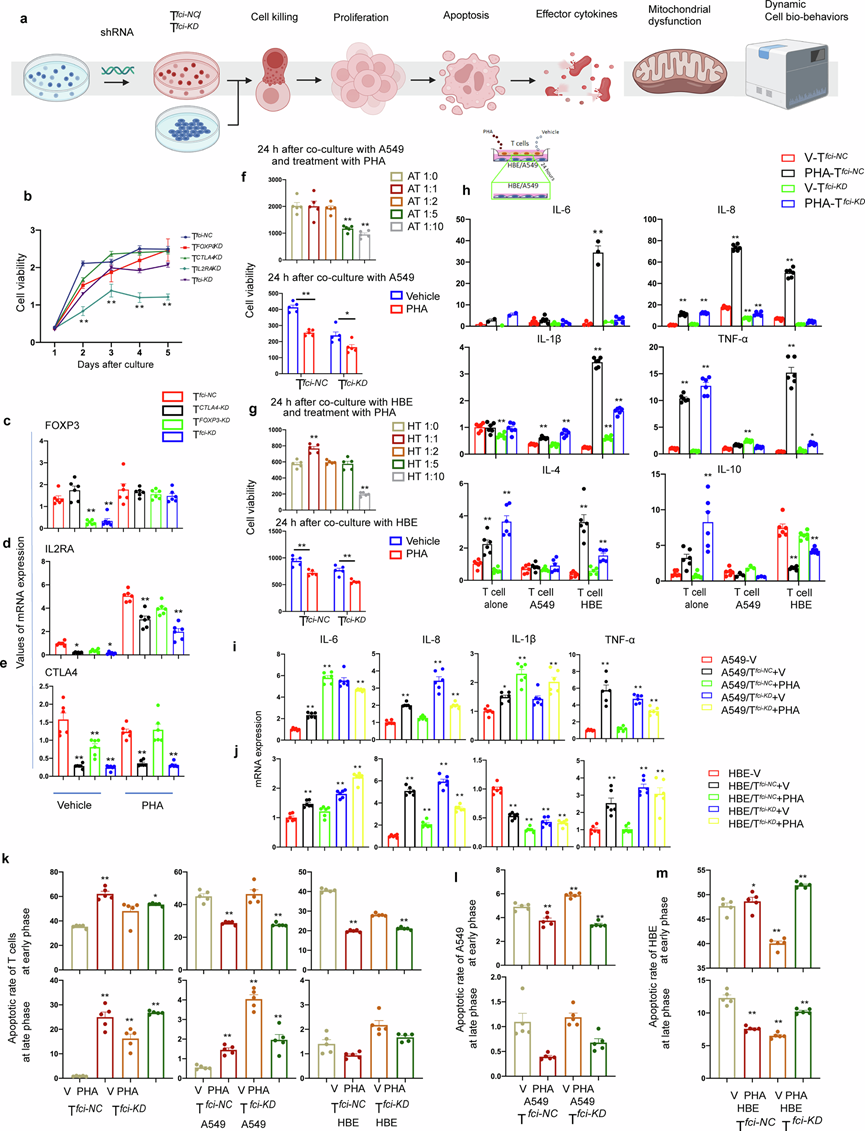
图5. Tfci-KD的功能表征及上皮细胞共培养反应。
(a) 实验流程图。(b) 细胞活力分析。(c-e) 细胞中FOXP3、CTLA4 和IL2RA 基因进行定量实时 PCR 验证。(f-g) 细胞活力分析。(h) 检测炎症细胞因子 IL-6、IL-8、IL-1β、TNFα 以及 Treg 相关细胞因子 IL-4 和 IL-10 的表达。(i) 检测 A549 细胞中 IL-6、IL-8、IL-1β 和 TNF-α 的基因表达。(j) 检测 HBE 细胞中 IL-6、IL-8、IL-1β 和 TNF-α 的基因表达。(k-m)流式细胞分析。
06
T 细胞转录组学分析和功能表征
为了评估 Tregfci ciMGP 对 T 细胞代谢和细胞行为的影响,他们检测了 ciMGP 敲低后的线粒体功能和生物能量学,发现 Tfci-KD 细胞的 ROS 生成水平较低,而与 HBE 细胞共培养则提高了经 PHA 刺激的 Tfci-KD和 Tfci-NC细胞的 ROS 水平。PHA 处理显著提高了 Tfci-KD细胞(无论是否与 A549/HBE 细胞共培养)的细胞内钙离子浓度,而 Tfci-NC细胞未观察到类似效应(图6a)。PHA 处理后,Tfci-NC细胞的线粒体质量增加,而 Tfci-KD细胞在载体或 PHA 处理下线粒体质量均降低(图6b)。HBE细胞和A549细胞的耗氧率 (OCR) 相当(图6c-d)。FCCP注射后,PHA处理的Tfci-KD组表现出最高的呼吸能力,而其他组的OCR水平相对较低(图6c)。在ROT/AA阶段,PHA处理的Tfci-KD组在共培养期间表现出最大的储备呼吸能力(图6d)。基于OCR的线粒体功能评估,包括基础呼吸、最大呼吸、非线粒体呼吸、ATP周转和质子泄漏,进一步揭示了ciMGP敲低诱导的生物能量重编程。Tfci -KD组的最大呼吸显著降低,基础呼吸和ATP周转增加,而PHA激活显著增强了Tfci-NC组的最大呼吸能力、基础呼吸和质子泄漏。为了揭示 Tregfci ciMGP 在细胞器间相互作用中的作用,他们评估了线粒体和内质网 (ER) 之间的形态和超微结构接触位点,发现 Tfci-KD比 Tfci-NC具有更多的线粒体-ER 接触,其中 PHA 刺激进一步增强了 Tfci-NC和 Tfci-KD中的这些接触(图6e)。
为了验证 Treg fci ciMGP与ETS1之间的相互作用,他们检测了NSCLC 组织中ETS1蛋白的定位,发现ETS1主要表达于淋巴细胞,并在癌细胞与邻近正常组织的交界处观察到核定位(图6l)。ETS1阳性细胞主要分布于 NSCLC 和邻近 NSCLC 组织的交界区域(图6l)。在正常肺组织中仅观察到少量 ETS1 阳性淋巴细胞(图 6l),而在 NSCLC 组织周围则出现较高密度的 ETS1 阳性淋巴细胞(图6l),并且偶尔会浸润到癌细胞微环境的中心和间质空间(图6l)。他们还定量分析了 T 细胞中 5 种ETS1家族同源物的表达。与载体处理组相比,PHA刺激显著上调了T fci -NC中的ETS1、ETV1和ETV4,以及Tfci-NC和Tfci-KD中的ETV6和ETS2 (图6m)。Tfci-KD中ETS1和ETV4水平显著降低,PHA处理后升高,其中ETV6表达水平高于PHA处理的T fci -NC 。为了阐明ETS1对Treg fci ciMGP的影响,通过过表达和敲低策略调控了T fci中的ETS1水平。研究发现,ETS1敲低降低了FOXP3和IL2RA的表达,并且当与T fci -KD联合使用时,所有3个靶基因的表达均进一步降低(图6n)。ETS1过表达增强了FOXP3和CTLA4的表达,但略微降低了IL2RA的表达(图6o)。ETS1沉默显著增加了线粒体-内质网接触位点的数量(图6p)。他们构建了一种条件性敲除小鼠模型,其中Ets1在表达Foxp3的细胞中特异性敲除(Ets1flox/floxFOXP3-Cre),并通过皮下接种Lewis肺癌(LLC)细胞建立了小鼠肺癌模型。
为了评估Ets1缺陷对Tregfci区室的影响,他们从荷瘤的Ets1flox/floxFOXP3-Cre小鼠及其Ets1 flox/flox对照组(每组n =6)中收集外周血白细胞(WBC)、脾脏和肺组织,并进行多参数流式细胞分析。结果显示,各组间WBC和肺组织中Treg fci的频率无显著差异。相比之下,Ets1flox/floxFOXP3-Cre小鼠脾脏中Treg fci的比例显著降低,表明 Tregfci对Ets1表达具有组织特异性依赖性。
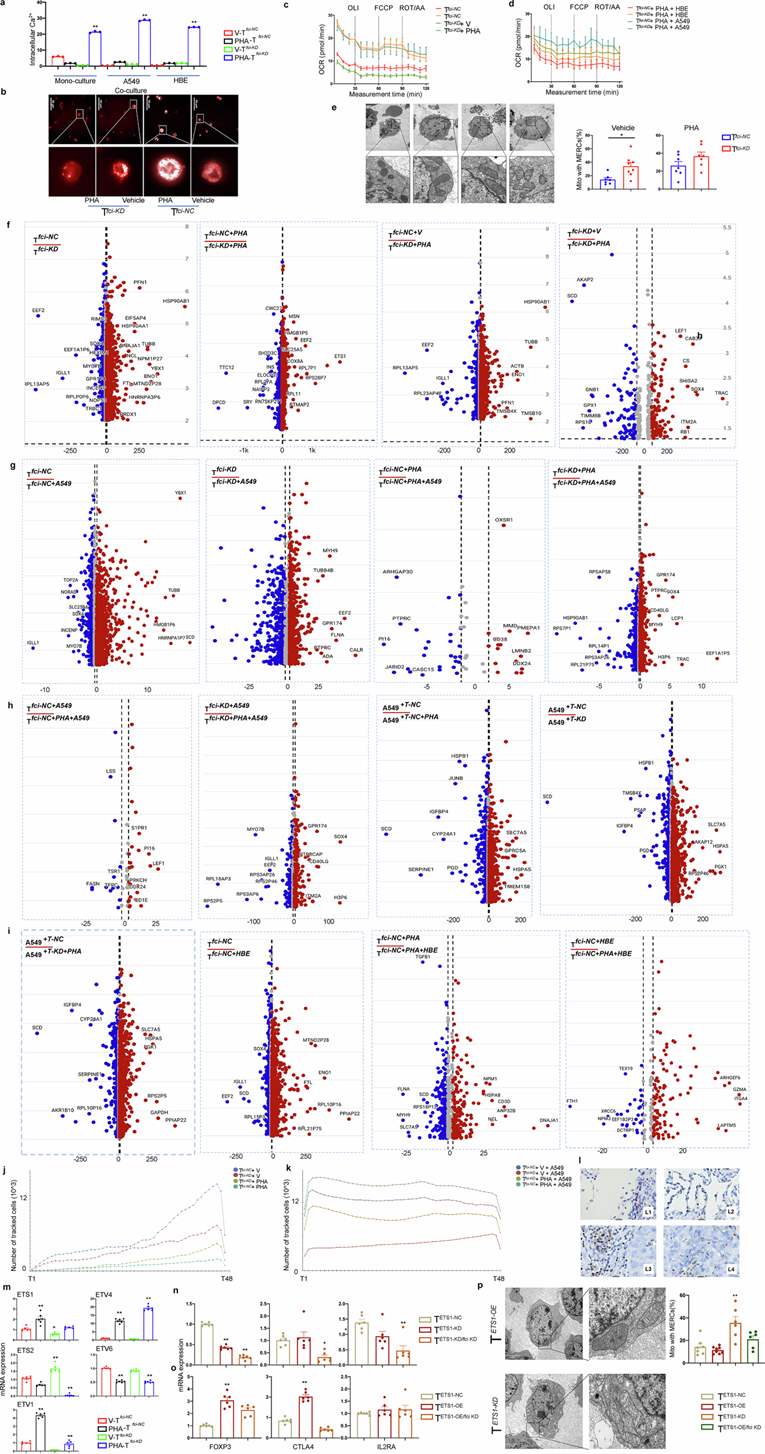
图6. 对T 细胞进行转录组学和功能表征。
(a) 代表性柱状图显示了使用 Fluo4 染料通过流式细胞技术测定的细胞内钙离子浓度。(b) 线粒体形态分析。(c-d) 细胞的耗氧率 (OCR) 。(e) 线粒体-内质网接触位点(MERC)的透射电镜(TEM)分析。(f-i) 转录组水平分析。(j-k) 细胞增殖情况分析。(l) 免疫组织化学染色。(m-o) mRNA 表达水平分析。(p) 使用透射电镜 (TEM) 分析。
07
单细胞转录组和代谢组分析
为了评估ETS1在Tregfci转录组谱中的调控作用,他们使用TK216抑制PBMC中ETS1蛋白的功能,并进行scRNA-seq。结果显示,PBMC中Tregfci的比例约为0.3%–0.4% (图7a)。与载体对照组相比,TK216处理组的Tregfci中约有98个基因显著上调,109个基因显著下调,并伴有细胞周期、IgA生成和代谢通路的变化(图7b-c)。为了评估ETS1在循环系统和肺组织中作用的异质性,他们使用ETS1条件性敲除小鼠(ETS1-cKO小鼠)或Ets1野生型小鼠(ETS1-wt小鼠)进行了额外研究,并基于ciMGP对小鼠血液白细胞和肺组织进行了scRNA-seq。循环系统中小鼠白细胞中Treg 的fci为 0.02–0.04%(图7d),与ETS1i-wt小鼠相比,ETS1-cKO小鼠血液中 759 个基因显著上调,85 个基因显著下调,涉及代谢和信号通路(图7e-f)。小鼠肺组织细胞中Treg细胞的fci百分比为0.26%–0.28%(图7g), ETS1-cKO小鼠肺组织中分别有49个或130个基因表达显著上调或下调。ETS1-cKO小鼠肺组织中前20个基因的表达谱与外周血中的表达谱不同(图7h),且ETS1 -cKO小鼠肺组织中代谢相关基因的主要变化显著降低(图7i)。在ETS1抑制的PBMC或小鼠血液和肺组织中,X染色体失活特异性转录本(XIST)、免疫球蛋白κ恒定区(IGKC)或DEAD-Box解旋酶3 Y连接区(DDX3Y)发生显著变化(图7j-k)。HLA -DRBS 和 PLCG2 在人样本中发生显著变化,而 MT-ATP6 和 MT-ND4 仅在小鼠样本中观察到。为了解这些因子在 17 个 Treg 亚群中的潜在特异性,基于 ciMGP 对所有 Treg 亚群进行了注释,并分析了其在PBMC中的表达。在ETS1抑制的人 PBMC(图7l)或ETS1-cKO小鼠的血液(图7m)和肺组织(图7n)中,Tregfci和 Treg -FOXP3的 IGKC mRNA 表达显著升高。在大多数含有 FOXP3、CTLA4 或 IL2RA 的 Treg 亚群中,XIST 或 DDX3Y mRNA 的表达上调或下调。
在明确Ets1在调控Tregfci转录组和代谢相关基因中的重要作用后,他们探究了Ets1缺失是否会影响Treg fci所在微环境的代谢。为了阐明Ets1在代谢中的关联和作用,他们检测了ETS1-cKO小鼠和ETS1-wt小鼠的循环和肺组织代谢组学特征。在小鼠肺组织(图7o)和血液(图7t)中分别鉴定出约465种和281种差异代谢物,其中肺组织(图7p-q)和血浆(图7u-v)中分别有333种和193种上调,132种和88种下调。变量的贡献以及代谢物在肺组织(图7p)和血液(图7u)中的积累的影响和解释力,能够清晰地区分ETS1-cKO小鼠和ETS1-wt小鼠。差异代谢物的热图显示,肺组织代谢组学谱的变化在组内具有高度一致性(图7q),而血浆代谢组学谱的变化则表现出更大的变异性(图7v)。前100个差异代谢物的Z评分值表明,血浆中差异代谢物变化的幅度(图7w)高于肺组织(图7r),且ETS1-cKO小鼠和ETS1-wt小鼠之间的代谢物差异显著。肺组织(图7s)和血浆(图7x)中差异代谢物富集通路高度重叠,包括中心碳代谢、三羧酸循环以及氨基酸和胆固醇代谢。
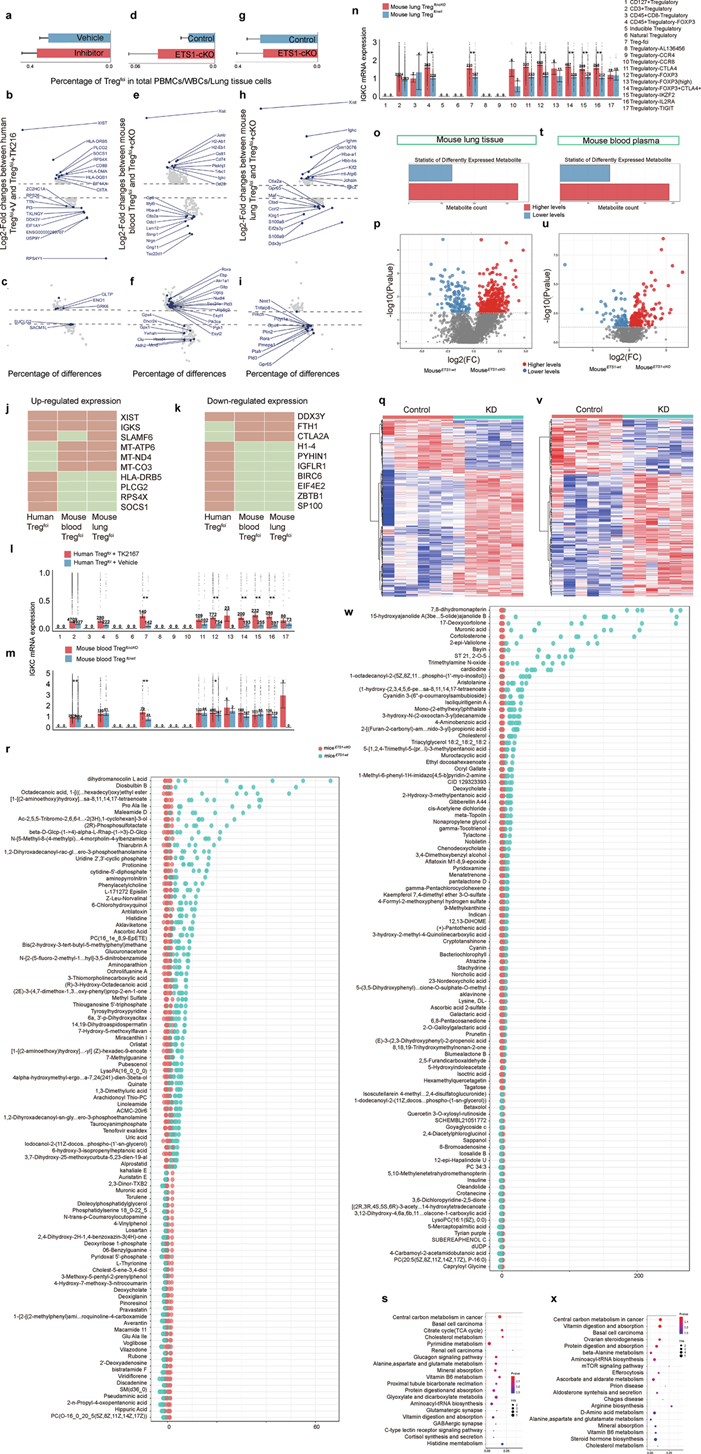
图7. 在ETS1抑制或缺失条件下,对人和小鼠的Tregfci进行单细胞转录组和代谢组分析。
(a, d, g) 计算Tregfci的百分比。(b-i) 表达差异显著的前20个基因或代谢相关基因的代表性变化。(j-x) 基因表达谱分析和代谢物分析。
+ + + + + + + + + + +
结 论
本研究利用多组学技术,鉴定并验证了一种Treg亚型(Tregfci),该亚型在空间、时间、疾病和预后方面均表现出疾病、器官和时间特异性特征。Tregfci在术前血液和NSCLC组织中显著富集,但在术后显著减少或消失。空间转录组和多重免疫染色显示,Tregfci主要定位于NSCLC与正常组织的交界处以及特定的微环境中。ETS1在 Tregfci中表达上调,并通过基因敲低和敲除实验得到证实。ETS1通过癌细胞来源的趋化作用促进 Tregfci从循环系统迁移至肿瘤组织。研究发现,ETS1与Tregfci 的ciMGP以及细胞内细胞器之间的相互作用能够调节代谢和线粒体功能。在用ETS1抑制剂处理的PBMC和ETS1-cKO小鼠的肺组织中,均证实了 ETS1 对Tregfci转录组和代谢谱的调控作用。此外,Tregfci和ETS1代表了 T 细胞独特的分子特征,是肺癌临床干预的潜在治疗靶点。
+ + + + +



