

 English
English文献解读|Cell Rep Med(14):多组学分析揭示肺肉瘤样癌的生物学和临床见解
✦ +
+
论文ID
原名:Multi-omics profiling unveils biological and clinical insights into pulmonary sarcomatoid carcinoma
译名:多组学分析揭示肺肉瘤样癌的生物学和临床见解
期刊:Cell Reports Medicine
影响因子:14
发表时间:2026.06.26
DOI号:10.1016/j.xcrm.2026.102887
背 景
肺肉瘤样癌(PSC)是一种罕见的非小细胞肺癌(NSCLC)组织学亚型,预后较差,约占所有肺癌的0.4%。世界卫生组织将PSC分为五种组织学亚型,包括多形性癌(PC)、梭形细胞癌(SCC)、巨细胞癌(GCC)、癌肉瘤和肺母细胞瘤,其中前三种亚型占PSC病例的90%以上。PSC的治疗遵循NSCLC的一线标准方案,但疗效并不理想,PSC表现出对常规治疗的耐药性,因此人们迫切需要新的治疗靶点和策略。免疫检查点阻断 (ICB) 和靶向治疗的免疫疗法显著改善了传统 NSCLC 的治疗,提高了患者的生存率,并有可能为 PSC 的治疗方法开辟途径。蛋白质在决定肿瘤表型方面至关重要,但迄今为止尚未有关于PSC的系统性蛋白质组学研究报道。
实验设计
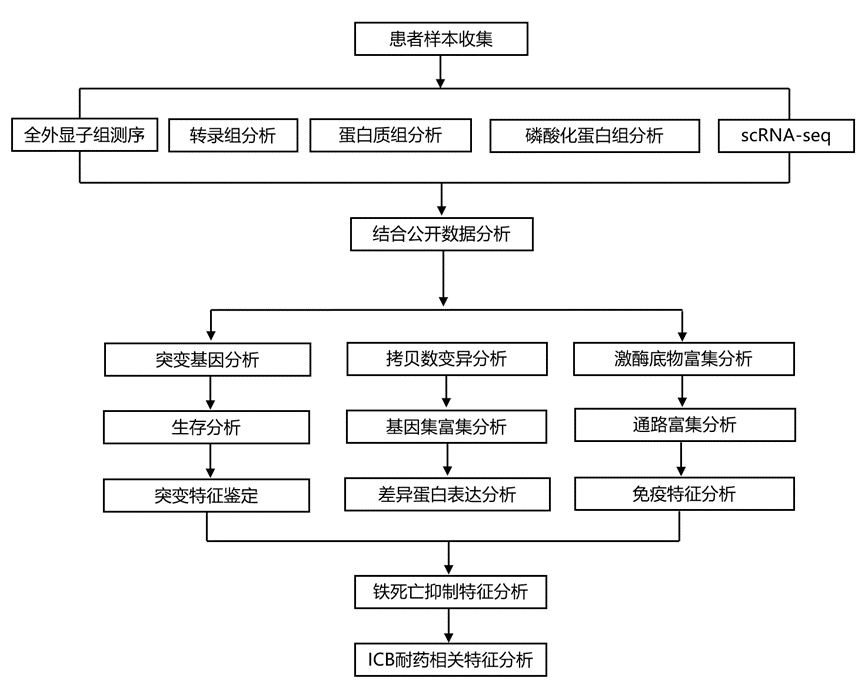
结 果
01

PSC蛋白质基因组图谱
为了表征PSC的蛋白质组学特征,研究团队收集了86例经手术切除的PSC患者的肿瘤组织及其配对的原发肿瘤组织样本(同济大学[TU]-PSC队列)。他们采用四种分子谱分析策略对TU-PSC队列进行表征,包括全外显子组测序(WES)、转录组分析(RNA-seq)以及基于同位素串联质谱标签(TMT)标记的蛋白质组学和磷酸化蛋白质组学分析(图1A)。利用蛋白质组学和磷酸化蛋白质组学数据,分别鉴定出9969个蛋白质和来自5514个磷酸化蛋白质的26002个可靠磷酸化位点(磷酸化位点)。整个数据集均保持了良好的重复性和较高的数据质量。样本相关性分析显示,与 NAT 样本相比,肿瘤样本中 mRNA 和蛋白质的表达一致性更高。
基于WES数据,他们评估了TU-PSC队列的基因组改变。基因组图谱基本证实了先前报道的PSC体细胞改变(图1B-C)。他们鉴定出TP53(65%)、KRAS(16%)、SMARCA4(12 %)、 PIK3CA(10%)和PTEN(7%)为显著的复发性体细胞突变(图1B)。TU-PSC队列的突变分布与其他三个PSC队列的突变分布大致相似。值得注意的是,TU-PSC队列中PTEN的突变率公开的研究结果相似。TU-PSC队列的KRAS突变率低于Nagano等人和Zhou等人的研究队列(图1C),提示可能存在地域差异。TU -PSC队列的KRAS突变率低于Nagano等人和Zhou等人队列,但与Yang等人的研究队列相当(图1C)。与既往PSC研究结果类似,TU-PSC队列中KRAS突变与较差的生存率相关(图1D)。他们发现肿瘤突变负荷(TMB)较高的患者总生存期(OS)更长(图1E),这与既往肺癌研究结果一致。TP53和SMARCA4等重要突变与TU - PSC队列中较高的TMB相关。
通过突变特征分析,他们在PSC中鉴定出三种主要的突变特征,即Sig1(Cosmic 4,吸烟相关突变)、Sig2(Cosmic 13,APOBEC相关突变)和Sig3(Cosmic 1,5-甲基胞嘧啶脱氨基相关突变)(图1F)。进一步分析表明,SMARCA4和KRAS突变患者的Sig1活性较高,而SMARCA4突变也与较低的Sig2活性相关。
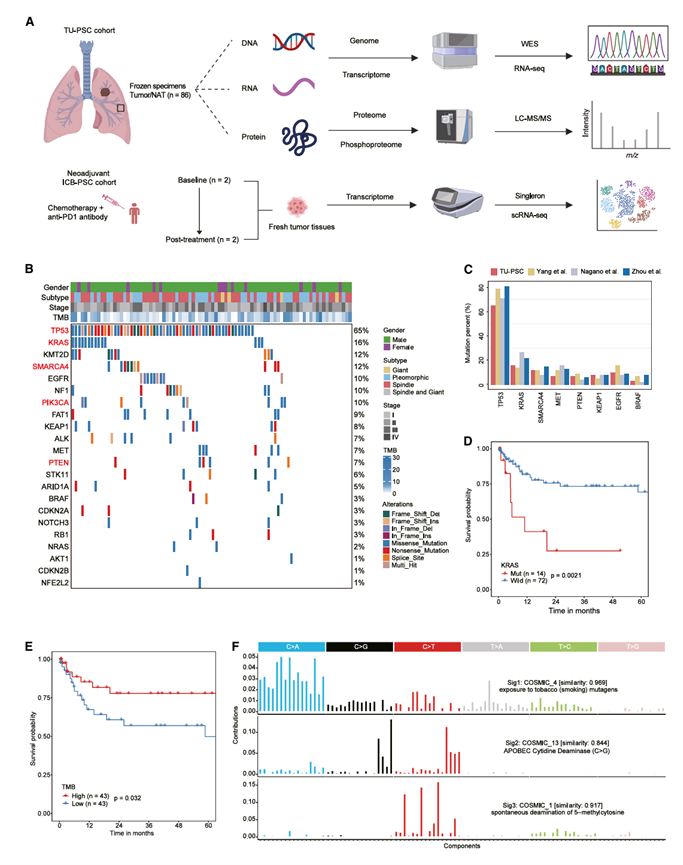
图1. PSC的多组学图谱。
(A) PSC多组学分析的工作流程。(B) TU-PSC队列的基因谱。(C) 四个PSC队列中体细胞突变的突变频率。(D) Kaplan-Meier 曲线比较KRAS突变型和KRAS野生型 (WT) 患者的总生存期。(E)Kaplan-Meier曲线比较高TMB和低TMB患者的总生存期。(F) TU-PSC队列中鉴定的三核苷酸基序频率图和富集突变特征。
02
基因组改变对PSC的影响
他们评估了体细胞拷贝数变异(CNA)对TU-PSC队列中mRNA、蛋白质和磷酸化蛋白表达的调控作用。WES分析鉴定出27,080个体细胞CNA,这些变异可能以“顺式”或“反式”模式影响mRNA、蛋白质和磷酸化蛋白的丰度。进一步聚焦于593个癌症相关基因(CAG),以确定CNA区域内的关键基因,其中590个CAG在TU-PSC队列中检测到(图2 A)。鉴定出 16 个 CAG 与所有三个组学水平上的显著顺式效应相关(图 2 A),这些 CAG 富集于 ErbB 信号通路、生长激素合成、分泌和作用、细菌侵袭上皮细胞、胰岛素信号通路和耶尔森氏菌感染(图 2 B)。
CNA分析发现TU-PSC队列中最常见的染色体扩增位于1q、5p、7p、7q和8q,而染色体8p的缺失最为常见(图2C)。此外,还发现驱动癌基因的扩增,包括EGFR(7p11.2,24例)、MALAT1(11q13.1,31例)、MYC(8q24,27例)、VEGFA(6p21.1,15例)和CDK6(7q21.2,22例),以及关键抑癌基因的缺失,例如CDKN2A(9p21.3,29例)(图2D)。对染色体臂水平体细胞CNA事件的生存分析显示,21p、13q、3p、16p、9q、Yp 和 Yq 的扩增与PSC的预后显著相关。位于 Yq 染色体上的 KDM5D 能够促进癌细胞的发生发展和转移。在 Yq 扩增的 PSC 肿瘤中,KDM5D 的表达显著升高(图 2 E)。
接下来,他们研究了PSC关键突变对其相应基因产物的基因组影响,并鉴定出五个具有显著顺式效应的基因(TP53、KRAS、PTEN、MET和SMARCA4 )(图2F-G)。KRAS突变肿瘤中KRAS的mRNA和蛋白表达均升高。同时,与非扩增肿瘤相比,KRAS扩增肿瘤中KRAS的蛋白和mRNA表达显著上调。此外,PTEN突变肿瘤中PTEN的蛋白表达降低(图2F)。MET扩增肿瘤中MET的蛋白和mRNA表达均升高,而MET突变与其蛋白或mRNA表达无相关性。SMARCA4是SWI/SNF染色质重塑复合物的ATP依赖性催化亚基,在SMARCA4突变肿瘤中,其mRNA和蛋白表达均降低(图2G)。此外,磷酸化蛋白质组学数据表明,SMARCA4突变肿瘤中SMARCA4上的六个磷酸化位点减少(图2H)。基于蛋白质组的基因集富集分析(GSEA)将SMARCA4突变与MYC靶基因、DNA复制和氧化磷酸化通路的上调(图2I)以及肌动蛋白细胞骨架调控、EMT和炎症反应通路的下调联系起来。值得注意的是,SMARCA4突变肿瘤中PRC2复合物的关键组分(包括EZH2、SUZ12和EED)的蛋白丰度显著升高(图2J)。SMARCA4突变预示着NSCLC患者对EZH2抑制剂敏感,提示EZH2抑制剂也可能成为SMARCA4突变型PSC的一种治疗选择。
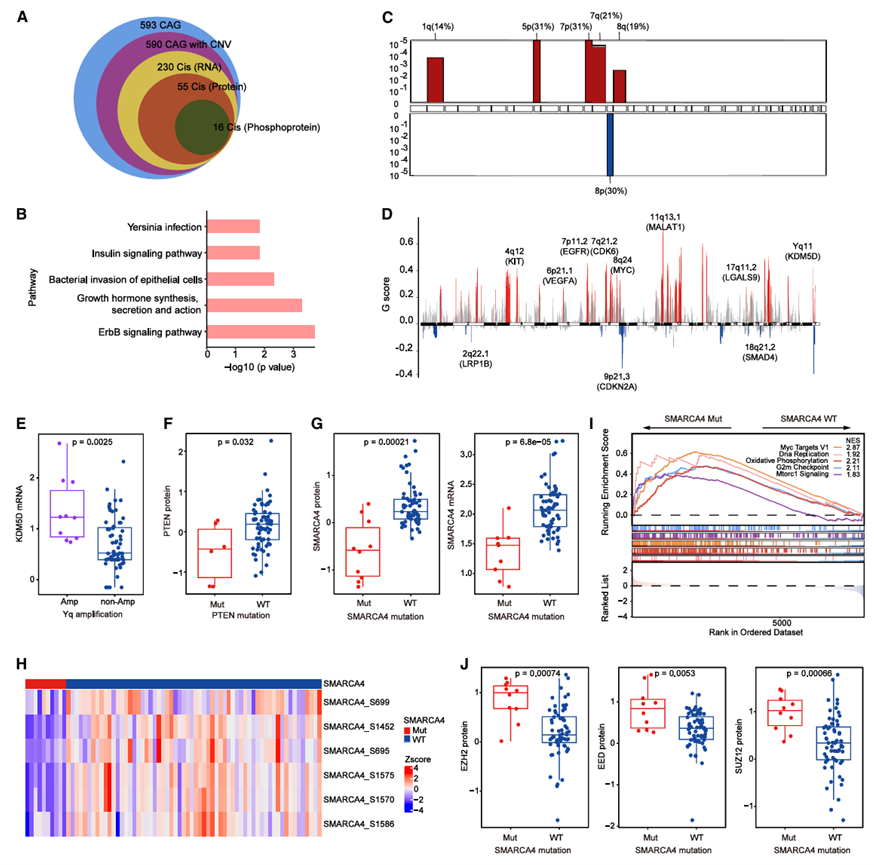
图2. 基因改变对PSC蛋白质组谱的影响。
(A) 维恩图显示了在多种数据类型中仅限于癌症相关基因(CAG)的显著顺式事件。(B) 在 mRNA、蛋白质和磷酸化蛋白质水平上共享的 16 个与顺式 CAG 相关的显著顺式事件的富集通路。(C) 全基因组臂扩增和缺失。(D) 全基因组局灶性扩增和缺失。(E) 箱线图显示 Yq 扩增肿瘤和 Yq 非扩增肿瘤之间 KDM5D mRNA 表达差异。(F) 箱线图显示PTEN突变型和PTEN野生型(WT)肿瘤之间 PTEN 蛋白的差异。(G) 箱线图显示SMARCA4突变型肿瘤和SMARCA4 WT 肿瘤之间 SMARCA4 表达差异。(H) SMARCA4突变型肿瘤和SMARCA4 WT 肿瘤之间 SMARCA4 磷酸化位点的热图。(I) 通过基因集富集分析发现,与SMARCA4 WT 肿瘤相比,SMARCA4突变肿瘤中显著上调的通路。(J) 箱线图显示SMARCA4突变型肿瘤和SMARCA4 WT 肿瘤之间 PRC2 复合物蛋白的差异。
03
与肿瘤发生和预后相关的蛋白质组学改变
随后,他们利用PSC肿瘤及其配对的癌旁组织(NAT)的蛋白质组学和磷酸化蛋白质组学谱,研究了PSC的蛋白质组学改变。主成分分析(PCA)显示,肿瘤和NAT在蛋白质和磷酸化蛋白质水平上均存在明显差异。鉴定出1681个差异表达蛋白,其中PSC肿瘤中847个蛋白表达上调,834个蛋白表达下调(图3 A)。GO富集分析显示,肿瘤中细胞质翻译、核糖体生物合成和rRNA加工相关基因表达上调,而伤口愈合、血液凝固和止血相关基因表达下调(图3 B)。应用更严格的差异表达标准,在肿瘤中鉴定出529个上调蛋白,将其命名为PSC相关蛋白(图3C)。利用人类蛋白质图谱(Human Protein Atlas)对这些蛋白的潜在临床应用价值进行了注释,该图谱包含230个血浆蛋白、124个酶、74个已知的癌症相关基因和44个已批准的药物靶点。在鉴定的PSC相关蛋白中,注意到SRSF5[一种富含丝氨酸/精氨酸(SR)的pre-mRNA剪接因子家族]在PSC肿瘤中蛋白表达增加,但mRNA表达降低(图3D)。SRSF5 在 mRNA 和蛋白质水平上相反的差异表达模式表明,对 TU-PSC 队列进行蛋白质组学分析对于指导后续 SRSF5 在 PSC 中的功能研究的实验设计至关重要。
磷酸化蛋白质组学数据分析鉴定出3221个差异表达的磷酸化位点,其中与配对的NAT相比,PSC肿瘤中1593个磷酸化位点上调,1628个磷酸化位点下调(图3E)。磷酸化位点与其对应蛋白之间的差异表达模式基本一致(图3F)。值得注意的是,1307个磷酸化位点的表达量比肿瘤中对应的蛋白显著增加(>2倍),尤其是在EMT通路VIM上的几个磷酸化位点(图3F)。他们利用激酶底物富集分析(KSEA)鉴定出26种在PSC肿瘤中活性显著增强的激酶(图3G),其中包括SRPK1,其在肿瘤中蛋白水平和磷酸化位点水平均升高(图3H)。观察到SRPK1蛋白表达水平较高的患者生存期更短(图3I),提示SRPK1在PSC中可能发挥致癌作用。
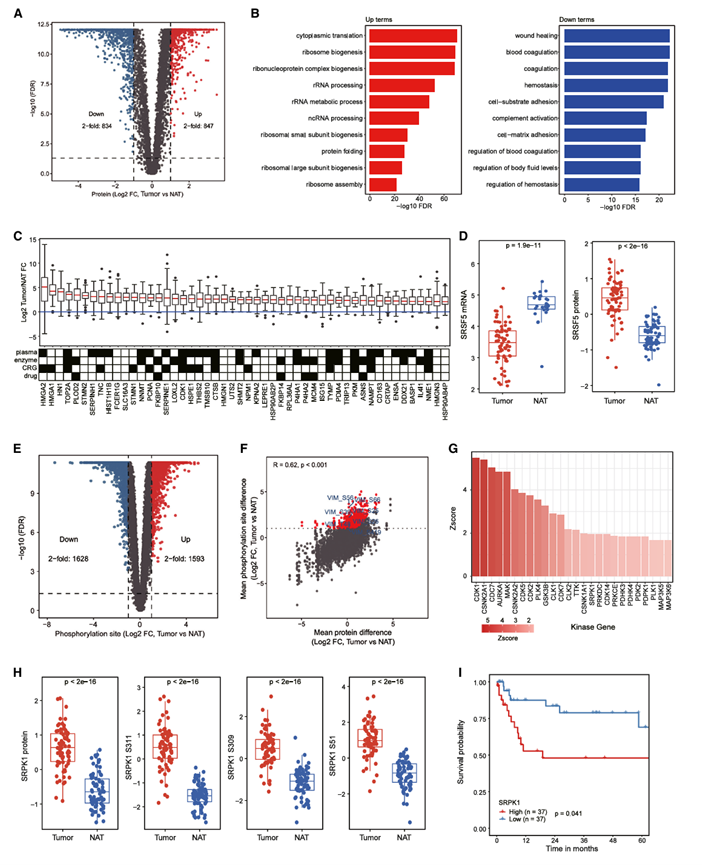
图3. 与肿瘤发生和预后相关的蛋白质组学和磷酸化蛋白质组学改变。
(A) 分析肿瘤和 NAT 之间的差异蛋白质表达。 (B) 与NAT相比,肿瘤中差异表达增加(左)和减少(右)的蛋白质富集的生物学过程。(C) 差异蛋白表达分析。(D) 箱线图显示肿瘤和NAT之间SRSF5 mRNA(左)和蛋白质(右)表达差异。(E) 检测肿瘤和 NAT 之间磷酸化位点水平的差异表达。(F) 比较肿瘤中蛋白质水平和磷酸化位点水平的丰度变化。(G) 肿瘤与NAT之间的激酶底物富集分析(KSEA)结果。(H) 肿瘤和NAT之间SRPK1蛋白和磷酸化位点的丰度差异。(I) 根据 SRPK1 蛋白丰度中位数比较总生存期的 Kaplan-Meier 曲线。
04
铁死亡的抑制促进了PSC细胞的增殖
在阐明PSC肿瘤及其配对的NAT之间的整体蛋白质组学变化后,他们尝试识别PSC中失调通路产生的潜在治疗靶点。EMT是PSC的标志性特征,间质表型对铁死亡的敏感性更高。铁死亡已成为肿瘤发生过程中的关键调控因子。有趣的是,对PSC肿瘤中847个显著上调蛋白的功能富集分析显示,铁死亡信号通路显著富集。值得注意的是,除CYBB外,其他五个上调的铁死亡相关基因(FTL、FTH1、PCBP1、PCBP2和SLC3A2)均注释为铁死亡抑制基因。在这五个铁死亡抑制基因中,SLC3A2和FTL与较差的生存率相关。与此一致的是,通过定量基于FerrDb数据库中注释的铁死亡抑制基因的铁死亡负调控(FNR)特征,这些观察结果表明PSC中铁死亡处于抑制状态。随后,研究了两种选定的铁死亡抑制因子SLC3A2和FTL在PSC中的潜在作用。SLC3A2和FTL在PSC肿瘤中均显著上调,且与TU-PSC队列中较差的生存率相关。在分别敲低SLC3A2和FTL后,评估了它们对PLA-801D细胞(一种人PSC细胞系)增殖的影响(图4 A-B)。结果表明,敲低SLC3A2和FTL后,细胞增殖显著降低(图4 B)。克隆形成实验也观察到了类似的结果(图4 C)。鉴于SLC3A2作为谷氨酸/胱氨酸反向转运蛋白的亚基所发挥的重要作用(该转运蛋白负责将胱氨酸与细胞内谷氨酸交换转运至细胞外),SLC3A2敲除后细胞培养上清液中细胞外谷胱甘肽(GSH)浓度显著降低(图4D)。FTL是铁蛋白的轻链亚基,对细胞内铁储存至关重要。使用FerroOrange荧光探针检测发现,FTL敲低后Fe 2+浓度显著升高(图4E)。此外,shFTL细胞中GPX4表达显著下调(图4F ),这与之前的研究结果一致。他们用DMSO或铁死亡抑制剂ferrostatin-1处理了不同组的PLA-801D细胞。用ferrostatin-1孵育后,shSLC3A2细胞的胞外GSH水平回升,同时GPX4蛋白表达上调(图4G-H)。ferrostatin-1处理后,shSLC3A2和shFTL细胞的增殖均部分恢复,而野生型(WT)细胞的增殖未受影响(图4I)。铁死亡通常与活性氧(ROS)的产生和脂质过氧化有关,因此他们评估了ROS水平和丙二醛(MDA)浓度(图4J-K)。沉默SLC3A2或FTL显著增加了ROS水平和MDA浓度,而抑制铁死亡后,二者均降低。除了MDA测定外,还使用C11 BODIPY评估了shSLC3A2和shFTL细胞中的脂质过氧化变化。当铁死亡增强时,C11 BODIPY 的丁二烯部分与活性氧 (ROS) 发生氧化还原反应,使其发射波长从 591 nm 偏移至 510 nm。利用流式细胞术检测了这种发射光谱的偏移,方法是测量异硫氰酸荧光素 (FITC) 和藻红蛋白 (PE) 通道之间的荧光强度比值。敲低 SLC3A2 或 FTL 后,大多数 PLA-801D 细胞的荧光强度比值显著升高,表明脂质过氧化和铁死亡水平升高。抑制铁死亡后,该比值恢复正常。综上所述,这些结果表明 SLC3A2 和 FTL 介导的铁死亡抑制可能促进PSC的进展,提示铁死亡可能是 PSC 的潜在治疗靶点(图 4 L)。
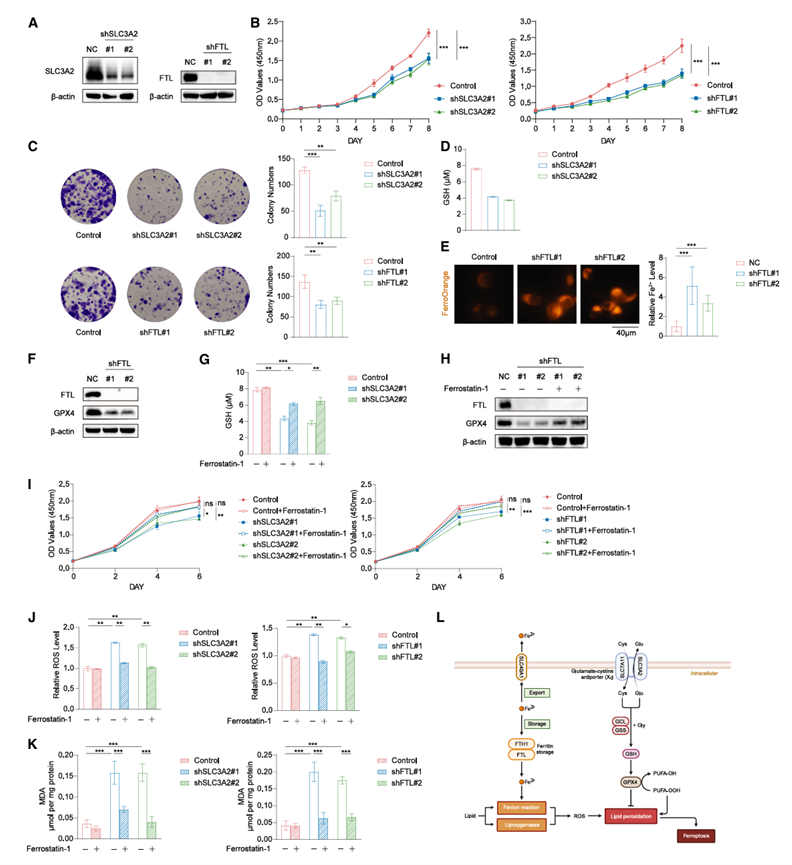
图4. 解除铁死亡抑制可降低 PSC 细胞增殖。
(A) SLC3A2和FTL在 PLA-801D 细胞中的敲低效率。(B) 分别在敲低SLC3A2(左)和FTL(右)后,使用 CCK-8 检测测量 PLA-801D 细胞的增殖情况。(C) 采用克隆形成实验检测PLA-801D细胞的增殖情况。(D) 在敲低SLC3A2后,检测 PLA-801D 细胞的细胞外 GSH 浓度。(E) 敲低FTL后,PLA-801D 细胞分别用 FerroOrange 染色,并使用微孔板读数仪进行定量。(F) 敲低FTL后,用指定的抗体对 PLA-801D 细胞的裂解物进行免疫印迹分析。(G) 敲低SLC3A2后,用 10 μM ferrostatin-1 处理 PLA-801D 细胞 24 小时。之后,检测细胞外 GSH 浓度。(H) 敲低FTL后,用 10 μM ferrostatin-1 处理 PLA-801D 细胞 24 小时。然后用指定的抗体对 PLA-801D 细胞裂解物进行免疫印迹分析。(I) 敲低SLC3A2(左)或FTL(右)后,用 DMSO 或 10 μM ferrostatin-1 处理 PLA-801D 细胞 24 小时。然后分别通过 CCK-8 检测 PLA-801D 细胞的增殖情况。(J-K) 敲低SLC3A2(左)或FTL(右)后,用 DMSO 或 10 μM ferrostatin-1 处理 PLA-801D 细胞 24 小时。然后分别向培养基中加入荧光探针以检测 ROS 的相对水平(J);收集细胞裂解液进行二喹啉甲酸测定和脂质过氧化 MDA 的检测(K)。(L) PSC中铁死亡抑制的图形模型。
05
PSC的免疫图谱
接下来,他们研究了PSC的免疫图谱。利用xCell衍生的细胞类型富集评分对肿瘤和NAT进行无监督共识聚类,识别出两组肿瘤,分别代表免疫学上的“热”聚类和“冷”聚类,以及一个NAT富集聚类(图5A)。与“冷”聚类相比,“热”聚类中免疫细胞(B细胞、CD4+T细胞、CD8+ T细胞和M1巨噬细胞)数量增加,免疫检查点表达增强,免疫评分升高,而肿瘤免疫功能障碍和排斥(TIDE)评分降低(图5A-C),表明免疫检查点阻断疗法(ICB)可能对“热”聚类有益。基因集富集分析(GSEA)结果显示,抗原加工和呈递(APP)、干扰素α、干扰素γ和T细胞受体(TCR)通路显著富集(图5D)。与热肿瘤聚类中APP通路富集相一致,该肿瘤聚类中也观察到更高的APP蛋白活性。
接下来,他们试图探究PSC免疫冷肿瘤中低免疫浸润的肿瘤内在决定因素。将组织学类型与免疫表型关联起来后,观察到冷肿瘤聚类中PC亚型的比例较高(图5E)。此外,PC亚型中干扰素α和干扰素γ通路显著下调。PC亚型中CD4 +记忆T细胞、树突状细胞(DC)和内皮细胞的浸润也较低。此外,PC亚型的免疫评分也低于SCC亚型。随后,他们探究了PSC中组织学亚型特异性突变,发现PC亚型中PTEN突变率相对较高。同时,所有PTEN突变型肿瘤均为冷肿瘤(图5F)。此外,PTEN突变型肿瘤的免疫评分较低(图5G),且多种免疫细胞(如CD+ T细胞、CD4+ Tem细胞和树突状细胞)的浸润减少(图5H)。GSEA还显示PTEN突变型肿瘤中干扰素α和APP通路下调,表明PTEN突变可能在PSC免疫原性丧失中发挥作用。
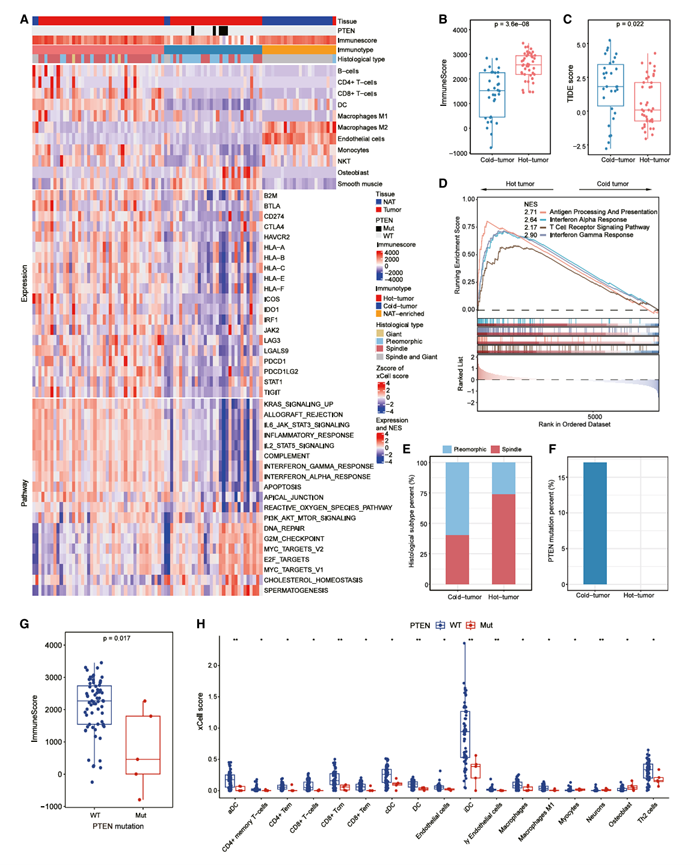
图5. PSC 的免疫图谱。
(A) 热图显示了三个免疫聚类中细胞类型组成、代表性免疫相关标志物的 mRNA 表达以及富集通路的活性。(B-C) 热肿瘤聚类和冷肿瘤聚类之间的免疫评分(B)和 TIDE 评分比较。(D) 通过基因集富集分析发现,与冷肿瘤相比,热肿瘤中蛋白质水平的通路显著激活。(E) 热肿瘤聚类和冷肿瘤聚类之间 PSC 组织学亚型分布的比较。(F) 热肿瘤聚类和冷肿瘤聚类之间PTEN突变率的比较。(G) PTEN突变型肿瘤与PTEN野生型(WT)肿瘤的免疫评分比较。(H) PTEN突变型肿瘤和PTEN野生型肿瘤免疫细胞xCell评分的比较。
06
PSC蛋白质组亚型和分子特征
接下来,他们分析了TU-PSC队列中的蛋白质组亚型。基于非负矩阵分解(NMF)的无监督聚类分析识别出三种不同的蛋白质组亚型,它们在基因组、免疫学和潜在治疗靶点方面均表现出独特的特征(图6A)。这三种蛋白质组亚型表现出不同的分子特征:S1亚型与蛋白酶体通路和较高的APP活性相关;S2亚型与EMT通路和最丰富的EMT相关磷酸化位点相关;S3亚型的特征是DNA复制通路富集和较高的PTEN突变率(图6A-C)。
蛋白质组学亚型与免疫表型的比较显示,S1 和 S2 亚型中以热肿瘤为主,而 S3 亚型中以冷肿瘤为主(图 6C)。与此一致的是,与 S3 亚型相比,S1 和 S2 亚型表现出更高的 PD-L1 表达(图 6D)。使用KEGG基因集进行分析显示,S1 亚型中与蛋白酶体和蛋白酶体复合物相关的基因富集(图 6E)。此外,S1 亚型还表现出多种蛋白酶体相关蛋白的更高表达水平。 PSMB8 和 PSMB9 是免疫蛋白酶体的两个亚基,它们与更高的抗原呈递水平以及非小细胞肺癌和黑色素瘤中更好的ICB反应相关。在S1 亚型中,PSMB8 和 PSMB9 特异性上调(图 6F-G),并且与PSC患者的生存期延长相关。与免疫蛋白酶体在抗原呈递中的重要作用相一致, APP 通路在 S1 亚型中也显著激活(图 6H-I)。
S2亚型肿瘤纯度相对较低,间质评分较高(图6J-K),表明该亚型癌成分较少,但肉瘤样特征较多。值得注意的是,EMT通路在S2亚型中特异性激活(图6B),这与基于RNA-seq数据在PSC肉瘤成分中观察到的较高间质特征一致。同样,EMT通路的关键组分在S2亚型中也表现出更高的表达。为了进一步证实这一结果,分析EM评分(数值越高表示肉瘤样特征越强,数值越低表示癌成分越多),发现S2亚型在mRNA和蛋白水平上的EM评分均较高(图6L-M),再次表明S2亚型具有更明显的肉瘤样特征。 TGF-β 是一种公认的强效 EMT 诱导剂,S2 亚型表现出显著激活的 TGF-β 信号通路。有趣的是,VIM 和 ZEB2(两种间质表型标志物)上的多个磷酸化位点在 S2 亚型中比相应的蛋白表现出更明显的诱导作用(图 6 N)。
DNA复制和细胞周期是S3亚型的标志性特征(图6B)。与其他两个亚型相比,观察到S3亚型具有更高的肿瘤纯度、更低的基质评分和更低的EM评分(图6J -M),这表明S3亚型在一定程度上具有更明显的癌特征。S3亚型显示出更高的经典LUSC标志物(SOX2和TP63)表达和更低的经典LUAD标志物(NAPSA和KRT7)表达。将体细胞突变与蛋白质组学亚型关联起来时,他们发现S3亚型中PTEN的突变率更高且PTEN蛋白表达下调,这可能导致S3亚型的免疫活性较低(图6C)。基于整合网络细胞特征库 (LINCS),他们筛选了能够逆转 S3 亚型特征的化合物,结果显示 S3 亚型中拓扑异构酶抑制剂 (TOPi) 显著富集。值得注意的是,TOP 家族的多个成员,包括 TOP1、TOP2A 和 TOP2B,在 S3 亚型中蛋白水平最为丰富。进一步证实,与其他肿瘤亚型相比,S3 亚型对拓扑替康的敏感性评分显著更高。除上述特征外,S3 亚型还表现出更高的 MYCN mRNA 表达水平和更高的干性评分。S3 亚型中某些表观遗传靶点(如 KDM1A、HDAC1 和 HDAC2)的蛋白丰度较高,提示表观遗传疗法可能适用于该亚型。
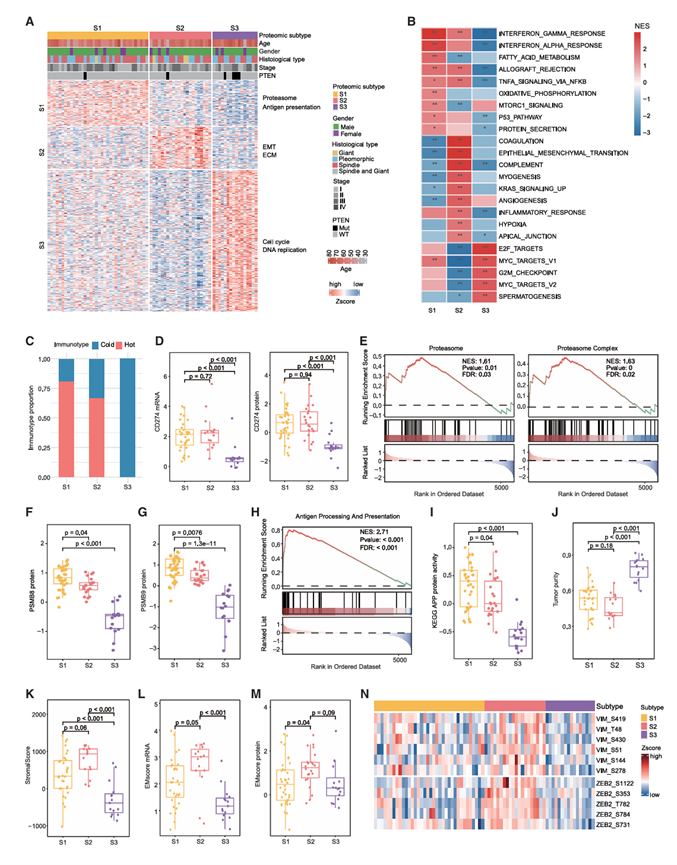
图6. TU-PSC 队列的蛋白质组亚型和分子特征。
(A) 热图显示了使用基于 NMF 的无监督聚类鉴定的三种蛋白质组亚型的临床病理特征、代表性富集的生物过程和特征蛋白。(B) 热图显示通过基因集富集分析在三种蛋白质组亚型中显著富集的通路。(C) 堆叠条形图显示每个蛋白质组亚型中 PSC 免疫类型(热肿瘤和冷肿瘤)的分布。(D) 比较三种蛋白质组亚型中 PD-L1 mRNA(左)和蛋白质(右)的表达。(E) 通过基因集富集分析发现 S1 亚型中蛋白酶体和蛋白酶体复合物通路显著富集。(F) 三种蛋白质组亚型中 PSMB8 蛋白表达的比较。(G) 比较三种蛋白质组亚型中 PSMB9 的蛋白质表达。(H) S1 亚型中抗原加工和呈递途径在蛋白质水平上显著富集。(I) 比较三种蛋白质组亚型的抗原加工和呈递途径蛋白活性。(J) 比较三种蛋白质组亚型的肿瘤纯度。(K)比较三种蛋白质组亚型之间的基质评分。(L-M) 比较三种蛋白质组亚型的 mRNA 和蛋白质。(N) S2 亚型中代表性 EMT 相关磷酸化位点的热图。
07
与PSC免疫检查点阻断疗法耐药相关的分子特征
临床上,PSC患者通常在晚期才确诊,新辅助免疫疗法的应用提高了NSCLC的手术切除率,可能也适用于PSC。免疫图谱和蛋白质组亚型分析表明,相当一部分具有热肿瘤行为的PSC患者可能从免疫检查点阻断(ICB)疗法中获益(图5A和图6A)。为了探索与PSC中ICB疗法疗效相关的分子特征,他们前瞻性地收集了3例接受新辅助化疗免疫治疗的PSC患者的4个新鲜PSC肿瘤样本,用于单细胞RNA测序(scRNA-seq)分析(新辅助ICB-PSC队列,图1A)。该队列包括两份治疗前活检样本,分别取自一名病理完全缓解(pCR)患者(组织1)和一名非主要病理缓解(NMPR)患者(组织4)(基线,n = 2),以及两份化疗免疫治疗后手术切除的样本(组织2,pCR;组织3,MPR)(治疗后,n = 2)。新辅助ICB-PSC队列还包括一名pCR患者的基线和治疗后配对样本(来自患者1的组织1和组织2)。去除双细胞并进行严格的质量控制后,他们获得了60932个细胞,并将这些细胞分为22个聚类,未观察到明显的批次效应,这些聚类主要由免疫细胞、上皮细胞和基质细胞组成。细胞聚类分析确定了12种主要细胞类型:B 细胞、树突状细胞、内皮细胞、上皮细胞、巨噬细胞、肥大细胞、单核细胞、中性粒细胞、自然杀伤 (NK) 细胞、浆细胞、基质细胞和 T 细胞(图 7 A),并通过对典型标记基因表达的手动注释予以证实(图 7 B)。
蛋白质组学亚型特征分析显示,S2亚型具有更高的基质评分和更明显的肉瘤样特征,并表现出EMT通路特异性富集(图6)。值得注意的是,已有研究报道EMT通路在原发性硬化性胆管炎(PSC)的癌变过程中发挥重要作用。因此,首先基于单细胞转录组分析(scRNA-seq)研究了EMT通路在PSC中从肺癌细胞向肉瘤样细胞转变过程中的作用。结合inferCNV的结果,将恶性细胞(癌细胞)归类于上皮细胞,将肉瘤样细胞归类于基质细胞和内皮细胞。进一步对所有scRNA数据中的恶性细胞进行了细胞轨迹分析。伪时间轨迹轴表明,EMT从起始区域的癌细胞转移到末端区域的肉瘤样细胞,提示EMT通路可能参与了PSC中癌细胞向肉瘤样细胞的转录重塑过程。伪时间表达动态分析进一步突出了从伪时间0到100逐渐上调的几个伪时间依赖性基因,包括参与EMT的COL1A1、COL6A1、TIMP1和VIM。与此一致的是,肉瘤样细胞中EMT相关基因和转录因子的表达高于癌细胞(图7 C-D)。除了EPCAM和KRT7表达较高外,癌细胞中多种主要组织相容性复合体(MHC)-II成员的表达也较高(图7 E-F)。随后,他们探讨了恶性细胞与免疫细胞之间的相互作用,结果显示,参与MHC-II通路(HLA-DMA_CD4、HLA-DMB_CD4、HLA-DQA1_CD4和HLA-DQB1_CD4)的配体-受体对在癌细胞与树突状细胞(DC)和巨噬细胞之间更为常见。然而,参与胶原相关通路(COL6A1_CD44、COL6A2_CD44和COL6A3_CD44)的配体-受体对在肉瘤样细胞与DC和巨噬细胞之间更为常见。
基于基线肿瘤细胞,GSEA分析显示NMPR患者中EMT相关通路富集(图7G)。与pCR患者相比,NMPR患者基线肿瘤中EMT相关基因(包括VIM、COL1A1、COL4A1和COL6A)高表达(图7H)。为了鉴定肿瘤间共有的细胞状态,虽然恶性细胞存在显著的患者特异性差异,使用Harmony算法将恶性细胞整合为6个肿瘤间共有的亚群。有趣的是,与pCR患者相比,NMPR患者肿瘤细胞中排名第一的C1程序活性更高,而其他五个程序则呈现相反的活性模式(图7I)。KEGG和GO富集分析显示C1程序中EMT相关过程富集(图7J)。这些结果表明,EMT通路激活可能与PSC中的ICB耐药性相关。值得注意的是,当将C1程序与TU-PSC队列中的蛋白质组亚型关联起来时,发现S2亚型的C1程序评分更高(图7K),这表明S1和S2蛋白质组亚型在TU-PSC队列中均表现出较高的PD-L1表达(图6D),但具有更高EMT特征的S2亚型可能对ICB免疫疗法耐药。
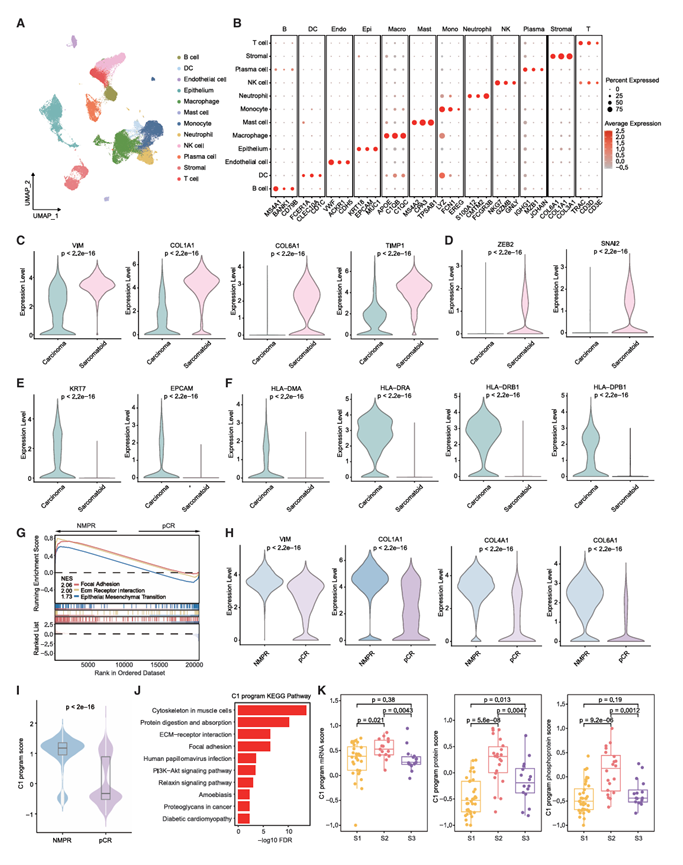
图7. 接受 ICB 免疫疗法的 PSC 患者的 scRNA-seq 图谱。
(A) 所有 scRNA-seq 数据的 UMAP 图,显示 12 种细胞类型。(B) 热图显示 12 种细胞类型中主要标记基因的表达。(C) 癌细胞和肉瘤样细胞之间 EMT 相关基因的表达比较。(D) 癌细胞和肉瘤样细胞中 EMT 相关转录因子的表达比较。(E) 癌细胞和肉瘤样细胞上皮基因的表达比较。(F) 比较癌细胞和肉瘤样细胞中 MHC-II 基因的表达。(G) 通过基因集富集分析,发现 NMPR 肿瘤与 pCR 肿瘤相比,其通路显著激活。(H) 基线样本中 NMPR 和 pCR 肿瘤之间 EMT 相关基因的表达比较。(I) 比较基线样本中 NMPR 和 pCR 肿瘤的 C1 程序评分。(J) C1 的显著富集 KEGG 通路。(K) 比较 TU-PSC 队列中三种蛋白质组亚型在 mRNA、蛋白质和磷酸化蛋白质水平上的 C1 程序评分。
+ + + + + + + + + + +
结 论
本研究整合了86例PSC患者的多组学数据,以及3例接受新辅助化疗免疫治疗患者的scRNA-seq数据,对PSC进行了多组学分析。他们发现PSC肿瘤中存在铁死亡抑制特征增强,并通过靶向FTL和SLC3A2证实了其对铁死亡的易感性。免疫图谱分析揭示了冷肿瘤、多形性癌和PTEN突变之间的关联。蛋白质组学分析鉴定出三种不同的患者亚型,每种亚型在微环境失调、基因改变和潜在治疗靶点方面均表现出独特的特征。上皮-间质转化在PSC中癌细胞向肉瘤样细胞的转化以及免疫治疗耐药性方面发挥着重要作用。总之,这种对PSC的多组学表征为深入研究其发病机制和识别潜在治疗靶点提供了丰富的资源。
+ + + + +



