

 English
English文献解读|Nature(48.5):小鼠视觉皮层发育过程中细胞类型的持续多样化
✦ +
+
论文ID
原名:Continuous cell-type diversification in mouse visual cortex development
译名:小鼠视觉皮层发育过程中细胞类型的持续多样化
期刊:Nature
影响因子:48.5
发表时间:2025.11.05
DOI号:10.1038/s41586-025-09644-1.
背 景
哺乳动物大脑皮层控制着多种灵活且受动机驱动的行为,并且在认知功能更高级的物种(包括人类)中显著扩张。该脑区一直是研究其所包含的各种细胞类型及其如何形成功能特异性神经回路的重要场所。皮层中的细胞类型可以根据多种细胞特性进行定义,包括基因表达、形态、生理、连接性或其各种组合。过去十年,单细胞转录组(scRNA-seq)技术提供了全面而详细的细胞类型分类,在成年大脑的每个皮层区域中定义了大约100种转录组细胞类型(T型),这些分类在不同区域和不同物种(例如,从小鼠到人类)之间具有显著的一致性。目前对哺乳动物皮层发育过程中涉及的机制和过程的理解仍存在诸多空白。例如,特定细胞类型何时确立、成年皮层中观察到的细胞类型在胚胎期确立的程度以及谱系分化如何发生等问题,目前仍不清楚。
实验设计
结 果
01

小鼠视觉皮层发育细胞类型图谱
研究团队利用scRNA-seq和单细胞核转录组分析(snRNA-seq)和单细胞核转座酶可及染色质测序(snATAC-seq)构建了小鼠视觉皮层发育过程中的两个数据集,利用scRNA-seq数据构建了转录组细胞类型图谱和发育轨迹图。然后,利用snATAC-seq数据重建了发育过程中表观遗传染色质可及性图谱。
研究团队首先构建了91个scRNA-seq文库,最终得到包含913297个单细胞的数据集。该scRNA-seq数据集密集覆盖了胚胎期和出生后 34 个时间点,从胚胎第 11.5 天 (E11.5) 到出生后第 28 天 (P28) 以及成年期 P56(图1a)。
为了构建成年细胞类型的发育轨迹,他们首先利用近期建立的成年小鼠全脑分类体系进行了标记转移。艾伦脑细胞-全脑(ABC-WMB)图谱作为成年期细胞的参考,用于在聚类水平上确定细胞类型。然后,他们通过从较年长的细胞到较年轻的同步年龄进行连续的细胞类型标记转移,将成年细胞类型标记传递给较年轻的细胞,涵盖所有出生后年龄。总体而言,在P20-28年龄组中,从成年ABC-WMB图谱中转移了35个P56谷氨酸能细胞聚类中的35个、61个P56 GABA能细胞聚类中的60个以及20个P56胶质细胞聚类中的16个的标记,从而几乎涵盖了成年小鼠视觉皮层的所有细胞类型多样性。
通过对成年和发育中小鼠视觉皮层进行高分辨率转录组细胞类型分类,展示了这些复杂的分子关系,并以树状图和均匀流形近似投影(UMAP)图的形式进行可视化(图1b-j)。该分类包含四个嵌套层级:15个类别、40个亚类别、148个聚类和714个亚聚类。它包含了公开报道的已知的发育中新皮层的神经元和非神经元细胞类别,以及在此发现的许多过渡细胞类型和亚型,还生成了一个包含6724个差异表达基因的列表,这些基因在所有聚类和亚聚类之间均存在差异。神经元细胞类型及其祖细胞构成发育图谱的很大一部分,共分为10类:神经上皮细胞(NEC)、Cajal-Retzius(CR)谷氨酸能(Glut)细胞、放射状胶质细胞(RG)、中间祖细胞(IP)、未成熟神经元(IMN)、非端脑内投射(nIT) Glut、端脑内投射(IT) Glut、CTX-CGE GABA、CTX-MGE GABA和CNU-MGE GABA(图1b-c)。这10类进一步细分为29个亚类、109个聚类和599个亚聚类。nIT Glut类包含四个主要的谷氨酸能亚类——L5 端脑外投射(ET)、L5 近端投射(NP)、L6 皮质丘脑投射(CT)和L6b——以及一个L6b/CT ENT亚类,该亚类主要存在于E17-P3阶段,根据定位结果,它属于内嗅皮层(图1b-d)。IT Glut 类包含四个主要亚类——L2/3 IT、L4/5 IT、L5 IT 和 L6 IT——以及一个 CLA-EPd-CTX Car3 亚类,该亚类由与屏状核和内梨状核共有的独特 L6 细胞类型组成。
皮质GABA能神经元起源于三个皮层下祖细胞区:尾侧神经节隆起(CGE)、内侧神经节隆起(MGE)和视前区(POA)。这些区域的神经祖细胞产生IMN,IMN迁移至皮质并在那里成熟。数据显示,MGE GABA IMN分化为四个亚类——CTX-MGE类中的Sst Gaba、Pvalb Gaba、Pvalb chandelier Gaba和Lamp5 Lhx6 Gaba,以及一个亚类——CNU-MGE类中的Sst Chodl Gaba (图1b-d)。CGE GABA IMN逐渐分化为Vip Gaba、Sncg Gaba和Lamp5 Gaba亚类。
所有非神经元细胞类型分为5类——胶质母细胞、少突胶质前体细胞(OPC-Oligo)、星形胶质细胞(Astro-Epen)、免疫细胞和血管细胞——每类又进一步细分为11个亚类(图1b-d)。胶质母细胞是OPC-Oligo和星形胶质细胞的主要祖细胞类型之一。OPC-Oligo类包含两个亚类:少突胶质前体细胞(OPC,表达Olig1、Olig2和Pdgfra)和少突胶质细胞(Oligo,表达St18和Opalin)。星形胶质细胞(Astro-Epen)类包含一个端脑星形胶质细胞亚类:星形胶质细胞(Astro-TE,表达Apoe、Aqp4、Aldh1l1和Slc1a3)。免疫细胞类包含两个亚类:小胶质细胞(表达Siglech、Sall1和Ifitm10)和BAM(表达F13a1、Pf4和Mrc1)。血管细胞类别由五个亚类组成:ABC(Slc47a1)、VLMC(Col1a1、Col1a2、Apod和Slc6a13)、周细胞(Kcnj8)、SMC(Acta2和Myh11)和内皮细胞(Endo,表达Ly6c1和Slco1a4)。
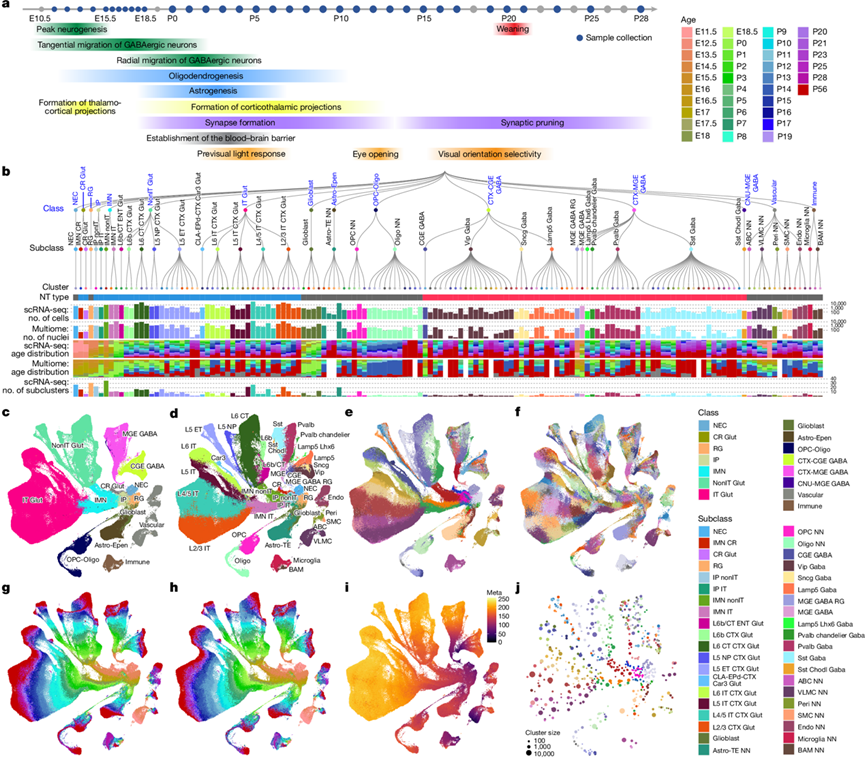
图1. 小鼠视觉皮层的转录组发育细胞类型图谱。
(a) 时间线示意图。(b) 148个聚类的转录组分类树状图。(c-i) 所有细胞类型的 UMAP 表示。(j) 星座图,显示按聚类着色。
02
构建细胞类型发育轨迹
轨迹分析是模拟细胞发育和分化动态过程的重要工具。鉴于成体阶段的细胞类型特征以及密集的时间采样,他们能够逐步传播相邻两个年龄之间的细胞类型特征。因此,他们基于同步年龄的整合空间,在出生后轨迹中定义了轨迹树的边权重,并采用了基于Monocle3的伪时间k近邻(k -NN)方法;而对于胚胎轨迹,则使用了基于Monocle3的伪时间的k-NN方法。为了简化发育过程的可视化和概念化,他们选择聚类与其一个前因之间权重最大的边,构建了从E11.5到P56整个时间线的发育轨迹图(图2a)。值得注意的是,用于构建所有轨迹的987条权重最大的边的平均权重为0.71,而未选择的321条边的权重小于0.5,平均权重为0.29。这一发现表明存在相对明确的发育轨迹模式。然后,他们根据整个发育轨迹图(图1i,图3a-d)分析了整体伪时间。
他们构建了视觉皮层中神经元和胶质细胞亚类的分支轨迹树(图2a)。该树与之前的研究基本一致,并得到了不同亚类相对比例随时间变化的趋势(图2b)以及每个分支节点的关键标记基因(图2c)的支持,其中UMAP图谱重点关注早期发育细胞类型(图2c)。轨迹树显示,最早从NEC中出现的细胞类型是IMN CR,它可能在E11.5之前起源于皮质边缘的NEC,并逐渐成熟为CR细胞。随后,RG分子特征在E13出现,紧接着在E13.5出现了IP nonIT和IMN nonIT分子特征。IP IT细胞和胶质母细胞以及IMN IT深层细胞的分子特征大约在胚胎发育第15.5天(E15.5)出现。IP非IT细胞分化为更多的IMN非IT细胞。IMN非IT细胞在胚胎发育第17天(E17)分化为三个亚类的非IT神经元(L5 ET、L6 CT和L6b),而第四个亚类(L5 NP)在胚胎发育第18.5天(E18.5)出现。IP IT细胞分化为更多的IMN IT细胞。IMN IT深层细胞在胚胎发育第17天(E17)分化为L6 IT和L5 IT神经元,而IMN IT上层细胞在胚胎发育第18.5天(E18.5)分化为L4/5 IT和L2/3 IT神经元。与此同时,胶质母细胞在胚胎发育第17天(E17)左右分化为星形胶质细胞和OPC。另外,对于 GABA 能神经元类别,MGE RG 和 MGE IMN 在 E11.5 之前出现,MGE IMN 在 E14.5 分化为 Sst 和 Pvalb 神经元。CGE IMN 在 E15.5 左右较晚出现在皮层中,并在 E18–P2 分化为 Vip、Sncg 和 Lamp5 神经元。
需要注意的是,细胞的身份是由其实时转录谱(分子身份)而非其出生日期定义的。在最早的阶段(E11.5-E12.5),源自大脑皮层的细胞主要由NEC(表达Hmga2)、CR细胞和早期产生的CR细胞(表达Ebf1、Ebf2、Ebf3、Reln、Calb2、Crabp2和Trp73)组成(分支节点1;图2-c)。IMN CR细胞是CR细胞的前体细胞,随着CR细胞的成熟,Eomes、Neurog2、Neurod1和Neurod2的表达逐渐降低。
从胚胎第13天(E13)开始,RG的分子特征(Sox2、Pax6、Hes1和Hes5)逐渐显现,RG同时产生IP(Eomes、Btg2、Neurog2和Gadd45g)和IMN(Dcx、Neurod1、Neurod2、Neurod6、Tubb3和Tbr1),这与直接和间接神经发生的共存相一致。E13.5至E16.5之间产生的大多数IP是non-IT细胞,并最终转化为non-IT细胞;而E17至P0之间产生的大多数IP是IT细胞,并最终转化为IT细胞(图2b)。虽然IMN IT深层细胞和非IT细胞都位于深层,但IMN IT深层细胞的出现时间晚于IMN非IT细胞(图2a-c)。
非IT的IP和IMN表达Neurog1、Lhx9、Fezf2、St18、Rmst、Nhlh1、Nhlh2和Kif26a,而IT的IP和IMN表达Pou3f1、Pou3f2、Pou3f3、Kif26b、Lama2和Slco1c1(节点3;图2a-c)。典型的深层神经元标记物Fezf2、Bcl11b、Foxp2和Tle4均在非IT细胞中选择性表达,但其表达的时间动态各不相同,其中Bcl11b和Fezf2在IP阶段出现,而Foxp2和Tle4则出现在IMN晚期。
IT和非IT轨迹之间的转录组差异不仅存在于IP和IMN中,而且在不同年龄的RG中也已存在,其中非IT细胞标记物Rmst存在于早期RG中,而IP标记物Pou3f2存在于晚期RG中(图2d)。最近的研究表明,皮质RG的转录谱会随着其生成非IT神经元、IT神经元和胶质细胞而发生变化。GABA神经元(nonIT Glut和IT Glut)和胶质细胞(OPC-Oligo和Astro)祖细胞的分化可能最早始于E15.5(节点2;图2a-c),并且RG亚类显示出不同转录组状态之间的连续性细胞(图2d)。首先,早期RG细胞富集Neurog2和Tenm4,这可能代表一种已确定的神经发生状态;而Tnc的表达则见于晚期RG细胞,这可能代表一种已确定的胶质细胞发生状态。其次,胶质母细胞在E15.5出现,并且Fabp7、Lipg、Slco1c1、Tnc、Qk和Slc1a3的表达水平高于RG细胞,这表明它们正在向胶质细胞方向转变。RG细胞已经表现出复杂的基因表达时间变化,并且它们在不同年龄阶段带着这些时间特征退出RG状态,分化为IP细胞、IMN细胞或胶质母细胞,最终分化为不同的神经元(nonIT和IT)或胶质细胞。这些结果与之前的谱系追踪和转录组分析研究中观察到的异质性一致,并可能解释了这种异质性。
他们利用近期生成的覆盖小鼠出生后第0天(P0)全脑的MERFISH数据集,确定了P0(一个关键的过渡时间点)发育细胞聚类的相对丰度和空间位置(图2e)。P0 MERFISH数据揭示了胚胎细胞类型以及持续到成年的新兴亚类的空间组织。此时,IP nonIT、IMN nonIT和IMN IT深层细胞数量稀少,而SVZ中仍存在大量IP IT细胞群,且IMN IT上层细胞广泛分布于整个皮层深度。这些结果与scRNA-seq结果一致(图2a-c)。值得注意的是,IMN IT上层细胞聚类中的亚聚类可分为三组,各组具有不同的层分布模式,表明其持续进行放射状迁移。亚聚类1-2集中在SVZ内或附近,而亚聚类6-11则位于皮层表面附近。亚聚类3-5分布于其他两组之间的皮层深度范围内。这些空间分布模式与UMAP中亚聚类从不成熟状态到成熟状态的相对位置相对应。
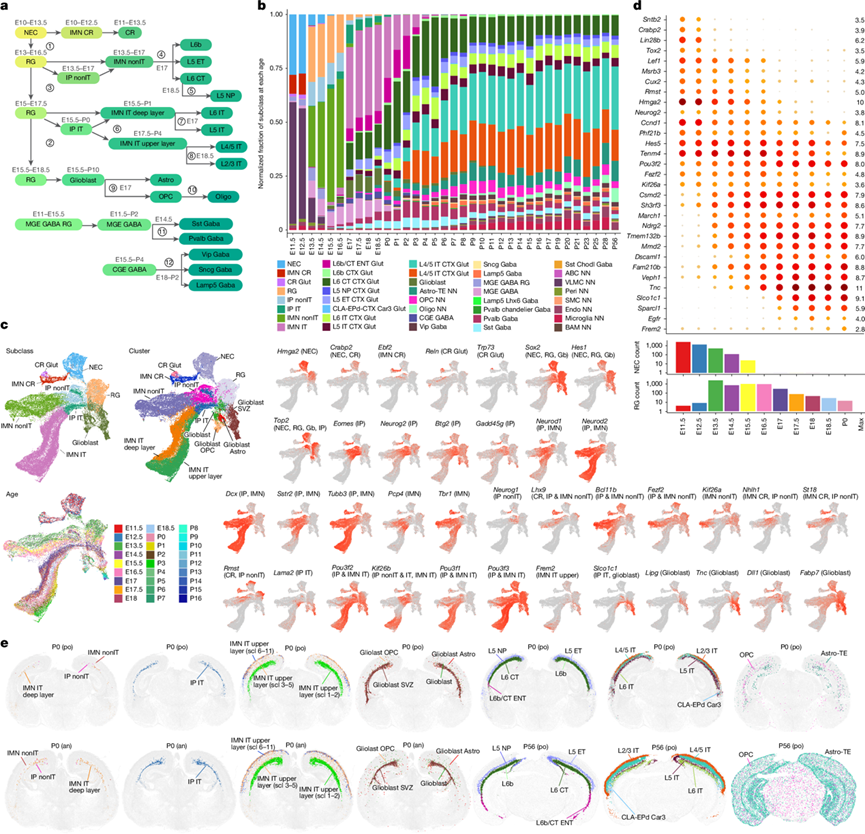
图2. 视觉皮层细胞亚类的发育轨迹。
(a) 视觉皮层细胞亚类的转录组轨迹,并估计了起始时间和主要分支节点。(b) 各年龄段不同亚类细胞的相对比例。(c) 早期发育细胞类型的UMAP图,按亚类、聚类、年龄和区分不同轨迹的关键标记基因的表达进行着色。(d) 散点图显示了NEC和RG细胞群中DE基因在胚胎发育阶段和P0时的表达情况。(e) P0和P56的代表性MERFISH切片,并标记了特定细胞类型。
03
谷氨酸能类型的发育轨迹
有丝分裂后的IMN细胞(IMN nonIT和IMN IT)逐渐分化为更多不同的细胞亚类和类型(图1c-f,图2a,图3a-e)。在nonIT细胞的发育轨迹中,IMN的分子特征(Fezf2、Bcl11b和Neurod2)在E13.5出现,并在IMN晚期阶段Foxp2、Tle4和Crym的表达逐渐增强。该发育轨迹在E17左右分裂为L6 CT、L5 ET和L6b(图2a-c)。晚期IMN nonIT细胞的基因表达谱与nonIT组中最常见的亚类L6 CT的基因表达谱非常相似。在L5 ET亚类中,Foxp2和Tle4表达下调,而Pou3f1和Bhlhe22表达上调。
在每个谷氨酸能亚类中,细胞持续分化和多样化,产生新的细胞聚类。他们构建了所有细胞类型的聚类轨迹树,并在每个分支点进行了差异表达基因分析(图3a-e)。在非IT类中,大多数L5 ET、L5 NP、L6 CT和L6b聚类在出生后第3天(P3)开始分化,但L5 ET聚类371-373(Chrna6)除外,它们代表了最独特的子集,并在关键期开始时分化(图3a-e)。
在IT细胞的演化轨迹中,许多早期分化的细胞聚类具有独特的层分布(图3a-e),并且许多具有独特层分布的基因在IT细胞类型分化的早期阶段表现出特定的表达模式。这一发现表明,皮层具有更精细的亚层梯度,这些梯度由出生后的早期阶段决定。在睁眼后的发育后期阶段,出现了更多的细胞聚类,这些新出现的细胞聚类与同源细胞聚类的空间分布差异较小。例如,L2/3层IT细胞聚类109在出生后第11天左右与细胞聚类110分化,表现为Bdnf表达增加和Adamts2表达减少。相比之下,细胞聚类118在出生后第21天进一步与细胞聚类109分化,表现为Baz1a和Tnfaip6表达增加。在L2/3层,细胞聚类118和110在出生后第56天的位置比细胞聚类109更浅。近期研究表明,体感皮层和视觉皮层中,L2/3 IT聚类在功能和发育上存在差异。他们还观察到L4/5 IT、L5 IT和L6 IT聚类的后期分化。在L4/5 IT中,聚类100是优势聚类,且为L4特有;而聚类101和聚类82分别在出生后第14天(P14)和第20天(P20)从聚类100分化而来。
从胚胎第15.5天(E15.5)开始,RG逐渐向胶质细胞生成转变,这表现为Tnc表达的增加(节点2;图2a-d)。在此期间,Slco1c1和Sparcl1在胚胎第17天(E17)于RG中激活,并在胶质母细胞中进一步激活。胶质母细胞的分子特征在E15.5出现,包含两个初始细胞聚类:胶质母细胞和称之为胶质母细胞SVZ的特殊细胞群(图3b,f)。胶质母细胞SVZ聚类与RG细胞共通表达Veph1、Tspan18、Tfap2c和Adamts18,并与星形胶质细胞共同表达Slco1c1和Tnc。Gja1在该细胞群中于出生后第0天左右开始表达,Thbs4于出生后第9天左右开始表达。
在出生后发育过程中,OPC占主导地位,但在出生后第11天(P11)之后,其比例逐渐下降。定向少突胶质细胞前体细胞(COP)大约在出生后第2天(P2)开始出现,其标志是Creb5、Etv5、Etv4、Sox9和Pdgfra的表达下调以及St18、Bmp4、Enpp6和Plp1的表达上调(节点10,图3b-f)。新生少突胶质细胞(NFOL)大约在出生后第11天(P11)出现,而成熟少突胶质细胞(MOL)大约在出生后第12天(P12)出现,其比例持续增加直至成年。这些结果与之前的研究一致,表明神经元活动会影响OPC的增殖、分化和髓鞘重塑。
在胚胎发育第11.5天(E11.5),最早出现的GABA能细胞群表达转录因子基因Dlx1、Dlx2、Ascl1和Gsx2,这些基因是底盖区所有GABA能神经元分化所必需的。
在胚胎发育第11.5天(E11.5),他们观察到内侧神经节隆起(MGE)GABA能祖细胞(即MGE GABA RG)的初步出现,这些祖细胞在胚胎发育第14.5天(E14.5)分化为MGE GABA IMN(图2a-b)。虽然之前的研究表明Sst和Pvalb细胞可能起源于MGE的不同区域,但他们并未在MGE RG和MGE亚类中观察到这两个细胞群的分离,尽管可能存在细微的差异基因表达。从E14.5开始,MGE细胞分化为两个亚类:Sst Gaba(Shisa6、Pou3f3、Npas1和Tox)和Pvalb Gaba(Adamts17、Shisa9、Tafa2和Zfp804b)(节点11;图2a)。他们还发现了三个截然不同的MGE亚类:Sst Chodl和Pvalb chandelier亚类(大约在出生后第1天出现),以及Lamp5 Lhx6亚类(大约在出生后第5天出现)。这些亚类可能在到达皮层之前就已经与其他MGE细胞类型分化开来。
在 Pvalb 和 Sst 亚类中,他们分别鉴定出五条主要的发育轨迹(图3c-g)。这些 GABA 能细胞类型按发育轨迹分组。每个轨迹组中的细胞聚类通常在出生后晚期发生分裂,尤其是在睁眼期(约 P11)或关键期(约 P21),这表明细胞分化仍在继续。
在 Pvalb 亚类(图3c-g)中,五种 Pvalb MET 类型中,四种是位于不同层的快速放电篮状细胞(或具有相关形态的细胞),一种(Pvalb-MET-5)是chandelier类型(对应于 Pvalb chandelier聚类 733)。发育组 1(Gpr149)和组 2(Reln和Pdlim3)均对应于 Pvalb-MET-3 类型(位于 L5)。组3(Tpbg和Calb1)包含聚类 742 和 752,其中聚类742 对应于 Pvalb-MET-4(位于 L2/3),聚类752 在 P17 时从聚类742 分化而来,对应于 Sst-MET-2。第 4 组(Sema3e、St6galnac5和Ptprk)对应于 L6 层中的 Pvalb-MET-2。Th + Pvalb聚类 735 对应于 L6 层中的 Pvalb-MET-1。然而,聚类735 的发育轨迹似乎非常模糊,其最近的祖先是 Sst聚类758。这一结果与之前的发现一致,即 L6 层中的Th + Pvalb 细胞可能是 Pvalb 和 Sst 亚类之间的过渡类型。
在Vip亚类中,他们鉴定出五种主要的发育轨迹,每组中的细胞聚类通常在出生后晚期发生分裂,这一结果表明细胞分化仍在持续(图3d-h)。大多数Vip细胞聚类存在于L2/3层,但639号细胞聚类富集于深层。Vip-MET-1-5型代表L2/3-5层的双极细胞或双聚类细胞。第1-3组中的大多数细胞聚类对应于Vip-MET-4和Vip-MET-5。641号细胞聚类对应于Vip-MET-2,而663号细胞聚类和第4组中的660-662号细胞聚类均对应于Vip-MET-1型。第5组(Grin3a和Igfbp6)是一种高度独特的Vip类型,几乎没有Vip表达,也没有匹配的MET类型。Sncg亚类具有一条主要发育轨迹,包含三个聚类,均对应于Sncg-MET-1类型,这是CCK+篮状细胞的主要类型。在Lamp5亚类中,他们鉴定了四条主要的发育轨迹。第2-4组的聚类706、708、709和718均对应于Lamp5-MET-1类型,代表L1-5层神经胶质样细胞。
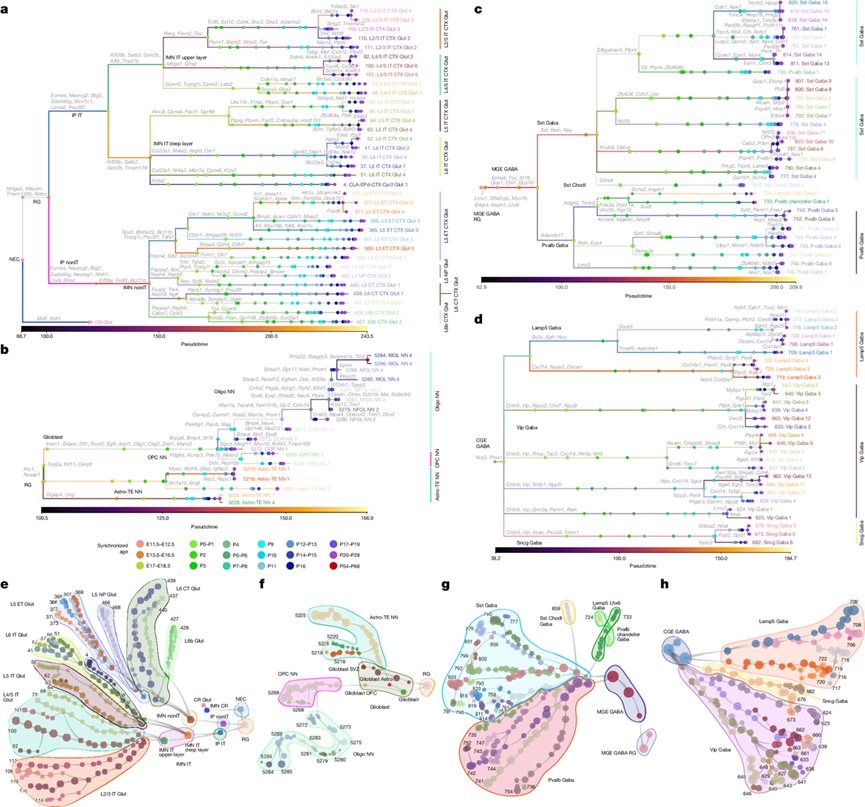
图3. 视觉皮层细胞类型的发育轨迹。
(a-h) 谷氨酸能、神经胶质、MGE和CGE细胞聚类的转录组轨迹树和星座图。
04
基因表达轨迹
为了分析发育过程中基因表达变化的动态过程,他们使用广义加性模型描述了每个亚类中4973个发育调控基因的时间变化曲线。将这些曲线聚类为36种轨迹模式,并进一步归类为5种一般发育基因轨迹:上调、瞬时上调、瞬时下调、下调和恒定(图4a)。在同一类别中,不同的模式可能存在细微但重要的差异。例如,轨迹2在胚胎早期上调,并在出生后早期达到平台期;而轨迹3在出生后上调,并在睁眼后达到平台期。
他们进一步根据基因在每个亚类中相似的时间轨迹对其进行聚类。虽然大多数基因在相关亚类中表现出相似的时间表达模式,但许多基因也显示出更微妙的亚类特异性时间动态。例如,虽然Tbr1的表达通常在神经发生过程中增加,但在IMN阶段之后,IT和非IT亚类中上层和下层神经元之间的Tbr1表达开始出现差异,尤其是在L5 ET细胞类型中显著下调(图4b)。许多重要的转录因子基因,包括Cux2、Foxp2、Nfia和Nfib,在不同亚类中表现出复杂的时间动态,这表明它们在皮层发育过程中具有严格调控的细胞类型特异性作用。
为了评估P56时转录因子和其他基因家族的表达与发育过程中转录组细胞类型的对应程度,他们鉴定了P56时每对亚类之间的差异表达基因(DEG)。利用这1035个成年DEG,以及3个精心整理的基因集(71个转录因子标记、139个功能基因和183个编码黏附分子的基因),训练了分类器来预测出生后不同年龄段(P0-P56)的亚类,并使用交叉验证评估了其性能(图4c)。正如预期的那样,亚类召回率的中位数在早期通常较低,但对于所有基因集,各个年龄段的亚类召回率仍然保持在中等到较高水平。召回率的波动在出生后早期更大,更多亚类的预测性能低于P56。这些发现突显了亚类在出生后发育过程中转录组的独特性,以及亚类身份即使在出生后早期也具有很强的稳健性。

图4. 发育过程中不同细胞类型和年龄的基因表达轨迹。
(a) 31个非恒定基因轨迹组的缩放表达量。(b) 选定标记基因的代表性基因表达轨迹。(c) 使用基于所有1035个成人标记基因(所有标记基因)、71个转录因子标记基因(转录因子基因)、139个功能标记基因(功能标记基因)和183个编码黏附分子的标记基因(黏附分子基因)构建的分类器。
05
睁眼期间的动态变化
上述轨迹分析揭示了视觉皮层细胞类型多样性从早期到晚期发育阶段逐渐增加,以及同一细胞类型内部广泛的转录异质性,如RG聚类所示(图2d)。为了定量多样性和异质性,他们绘制了同步发育阶段所有亚类的聚类和亚聚类总数(图5a)。聚类的数量随时间持续增加,并在P11-P13和P19-P21阶段出现跃升。相比之下,在亚聚类水平上,亚聚类数量出现了几次激增,这表明在与特定发育事件相关的不同时间段,过渡细胞亚型或细胞状态的异质性增强。这些事件包括神经发生(E13.5-P1)、轴突生长和突触形成(P5-P9)、睁眼(P12-P15)以及经验依赖性可塑性的关键期(P20-P28)。
为了定量出生后发育(P0-P28)期间不同细胞类型基因表达的动态变化,他们鉴定了每个亚类中相邻两个年龄组之间的DEG。在神经元细胞类型中,例如nonIT Glut、IT Glut和MGE GABA,在P0-P5期间以及睁眼前后(P10-P15)观察到显著的转录变化。此后,变化速率显著降低。非神经元细胞类型也表现出不同的模式。OPC-Oligo和星形胶质细胞-TE在P0-P4和P9-P13期间基因表达发生显著变化。睁眼后,OPC和星形胶质细胞在P14-P16期间出现显著变化,而少突胶质细胞在P17-P21和P25-P28期间发生显著变化。小胶质细胞在 P10-P12 和 P15-P17 期间表现出基因表达下调。
睁眼过程中发生的显著变化以及睁眼后新细胞类型的出现,促使他们探索睁眼前后分子特征的变化。他们首先对每个亚类或细胞聚类进行睁眼前后DEG分析,结合P7-P10(睁眼前)和P11-P15(睁眼后)的scRNA-seq数据(图5c-d)。值得注意的是,所有神经元和非神经元亚类均表现出不同的转录变化,每个亚类和细胞聚类中均存在上调或下调的基因(图5d)。平均而言,谷氨酸能神经元亚类(包括IT和nonIT亚类,每个亚类约有1200-2000个DEG)的DEG数量多于GABA能神经元亚类,Pvalb亚类除外。虽然大多数神经元亚类的上调基因数量多于下调基因数量,但所有非神经元亚类的下调基因数量均多于上调基因数量,尤其是小胶质细胞。

图5. 睁眼前后动态基因表达变化。
(a) 每个同步年龄的聚类和亚聚类的数量。(b) 按类别分组,定量发育过程中每个亚类中DEG的变化。(c) 神经元和非神经元类别的 UMAP。(d) 所有细胞亚类在睁眼前后不同年龄之间的DEG。
06
发育过程中染色质可及性的变化
通过整合 snRNA-seq 和 scRNA-seq 数据集(snMultiome),从参考 scRNA-seq 图谱中获得了每个细胞核的转移细胞类别、亚类别和聚类标签,他们将这些时间点合并成与同步年龄组一致的年龄组。基于整合的 scVI 潜在空间的 UMAP 显示 scRNA-seq 和 snRNA-seq 数据高度混合,并且两个数据集之间亚类别和年龄组的划分清晰一致(图6a-d)。
他们首先应用此方法分别研究了IT Glut和nonIT Glut亚类的特异性(图6e-f),因为许多基因在这两类中重复使用以指定不同的细胞类型。对于IT Glut亚类,他们确定了每个亚类的早期和晚期峰值基因对(图6e)。峰值模块1和2是IMN IT亚类特有的,并与Kif26b和Pou3f3等基因相关。峰值模块29和30对所有IT亚类的可及性逐渐增强,其中模块30的激活晚于模块29。Nr4a2 、Nr2f2和Car3及其对应的峰值与CLA-EPd-CTX Car3 Glut亚类相关,而Nr4a3和Col6a1与L6 IT相关。Fezf2、Etv1和Deptor与 L5 IT 相关联,Whrn和Pamr1与 L4/5 IT 相关联,Tpbg和Stard8与 L2/3 IT 相关联,而Pou3f2和Pou3f1则优先与上层相关联,且具有不同的时间动态。总体而言,细胞类型特异性转录因子倾向于早期激活,而其他功能基因则在后期激活。
对于nonIT Glut(图6f),Lhx2是IMN非IT亚类的特异性标记。对于L6b亚类,Hs3st3b1是早期标记,Moxd1和Cplx3在发育后期激活,而Nxph4的表达在发育过程中相对稳定。类似地,Syt6和Arhgap25分别是L6 CT的早期和晚期标记,Tshz2和Vw2cl分别是L5 NP的早期和晚期标记,Pou3f1和Lratd2分别是L5 ET的早期和晚期标记。这些标记基因都具有匹配的染色质可及性谱,并具有相似的亚类和时间特异性。此外,还有一些特异性的峰值模块,但多个亚类共有这些模块。例如,模块11是L6b和L6 CT共有的,模块20是L6 CT和L5 NP共有的,模块29是L5 NP和L5 ET共有的。在发育过程中通常上调的第27和28个峰值模块,在L5 NP中特异性地受到抑制。有趣的是,这些峰值模块在不同亚类之间的差异比IT Glut类的峰值模块更为显著,后者在不同亚类之间的差异则呈现更大的梯度变化。
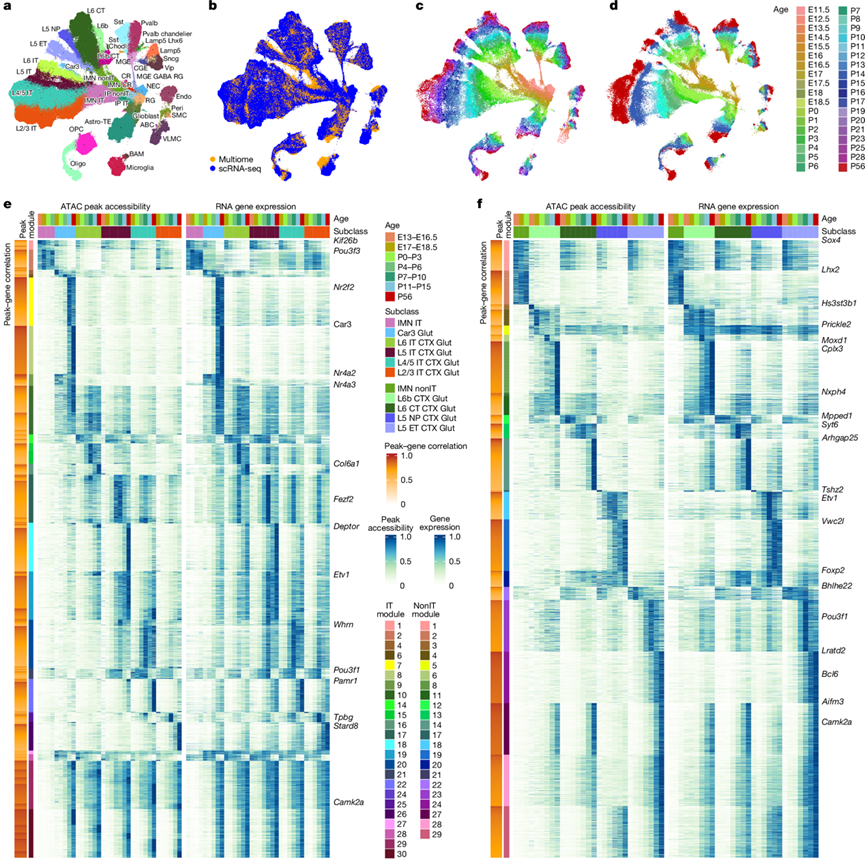
图6. snMultiome 数据的整合以及谷氨酸能神经元类型和发育过程中不同年龄段的染色质可及性和基因表达的对应关系。
(a-d) 整合空间中scRNA-seq细胞和snMultiome细胞核的UMAP。(e-f) IT亚类和nonIT亚类以及发育过程中不同年龄阶段染色质可及性和基因表达对应关系的热图表示。
07
转录因子调控因子和基因调控网络
为了鉴定每个峰模块的潜在转录因子(TF)调控因子,他们利用所有成对比较方法,对峰模块间的差异TF DNA结合基序进行了分析。对于在任何成对比较中均显著的TF基序,他们绘制了所有峰模块中该基序的出现频率,以及每个模块的平均亚类和时间可及性模式(图7a-c)。他们使用与差异TF基序相关的TF作为潜在调控因子,利用SCENIC+框架构建了基因调控网络(GRN)。他们的目标是识别激活和抑制性的TF相互作用。每个TF-峰-靶标三元组都赋予一个置信度评分,并且仅展示排名最高的预测结果。鉴于同一 TF 家族的基序通常表现出相似的富集模式,并且它们相关的 TF 可能具有很强的相关性,他们仅突出显示每个 TF 家族中最具影响力的调控因子,这是通过 GRN 分析确定的。
在IT Glut类神经元(图7a-b)中,Lhx2基序在IMN IT和L2/3 IT亚类特有的峰值模块中富集,而Lhx2表达在除L2/3层以外的IT神经元成熟过程中逐渐降低,这可能有助于维持上层神经元的特性。Nr4a2是CLA-EPd-CTX Car3亚类神经元的关键调控因子,其基序在Car3亚类神经元的早期和晚期特异性峰值模块中均富集。Rora和Rorb是L4/5 IT亚类神经元的关键调控因子,二者在L4/5 IT亚类神经元特异性峰值模块中均表现出最强的基序富集,并调控L4/5 IT亚类神经元的标记基因Whrn和Rspo1。Rora和Rorb具有高度相似的结合基序。Rora的表达范围更广,而Rorb则高度特异性地表达于L4/5 IT亚类神经元,它们可能协同调控同一组靶基因。POU-III类转录因子基因Pou3f1、Pou3f2和Pou3f3也是上层神经元的关键调控因子。这一发现与它们在决定和维持这些神经元群体的特性方面发挥的关键作用相一致,分析还预测Cux2是其下游靶点。
对于nonIT Glut类(图7c-d),他们鉴定出Nr4a2是L6b的调控因子,Ascl1和Etv1是L5 NP的调控因子,Pou3f1是L5 ET的调控因子,以及它们各自的细胞类型特异性靶点。与IT亚类类似,Bhlhe22和Neurod6在非IT亚类中具有相似的峰值模块富集,但表达模式相反。Bhlhe22和Neurod1在L5 ET中表达较强,而Neurod6表达较弱。分析表明,Bhlhe22可能通过与Neurod1的协同作用,抑制L5 ET中IMN非IT和L6 CT标记物的表达,同时激活L5 ET标记物的表达。
对IT和非IT亚类的GRN进行比较,发现几个共有调控因子(图7e):Car3和L6b中的Nr4a2,L5 IT和L5 NP中的Etv1,L2/3 IT和L5 ET中的Pou3f1,除L5 ET外,IT深层神经元和nonIT亚类中的Neurod6,以及IT上层神经元和L5 ET中的Neurod1和Bhlhe22。考虑到L5 ET是非IT类中最表层的神经元(图2e),这种TF可能反映了各亚类中神经元的相对层级位置。
除了调控细胞类型特异性的转录因子外,他们还鉴定出一个大多数皮层谷氨酸能细胞类型共有的保守时间调控网络(图7f-h)。Tcf4和Sox4在IP期上调,并在IMN期后逐渐下调。他们推测Sox4同时抑制某些神经元标记物的过早表达,特别是L6 CT标记物(图7c-d),以帮助维持细胞处于IMN状态。Mef2c和Mef2d在整个发育过程中持续增加,在神经元分化、突触连接和神经元存活中发挥着关键作用。
他们发现,包含Junb和Fos的AP-1复合物是睁眼后活动依赖性基因表达的核心调控因子(图7a-g)。AP-1受突触活动激活,调控其他早期基因(IEG),例如参与突触重塑和成熟的Arc。许多在成年期表达和染色质可及性达到峰值的基因可能是AP-1的靶基因。转录抑制因子Bach2是结合AP-1样基序的关键时间调控因子(图7f-h)。与Junb和Fos表达在成年期增加不同,Bach2的表达在睁眼后下降。Bach2通过竞争共有的结合位点来拮抗AP-1的活性,这与其在免疫系统中抑制AP-1驱动的转录的已知功能相一致。
此外,他们发现Hlf和Etv5在睁眼后均上调,是晚期神经元发育的重要调控因子(图7f-h)。Etv5参与轴突生长、树突分支和神经环路形成,在背根神经节和海马发育过程中受神经营养因子BDNF的调控,而BDNF本身在睁眼后也上调。综上所述,这些发现揭示了一个核心的时间转录网络,该网络协调皮层谷氨酸能神经元成熟的各个阶段。该网络平衡了早期神经发生、分化和突触发育与抑制过早的基因激活,最终确保了神经环路的正常形成、活动依赖性可塑性和长期的神经稳定性。
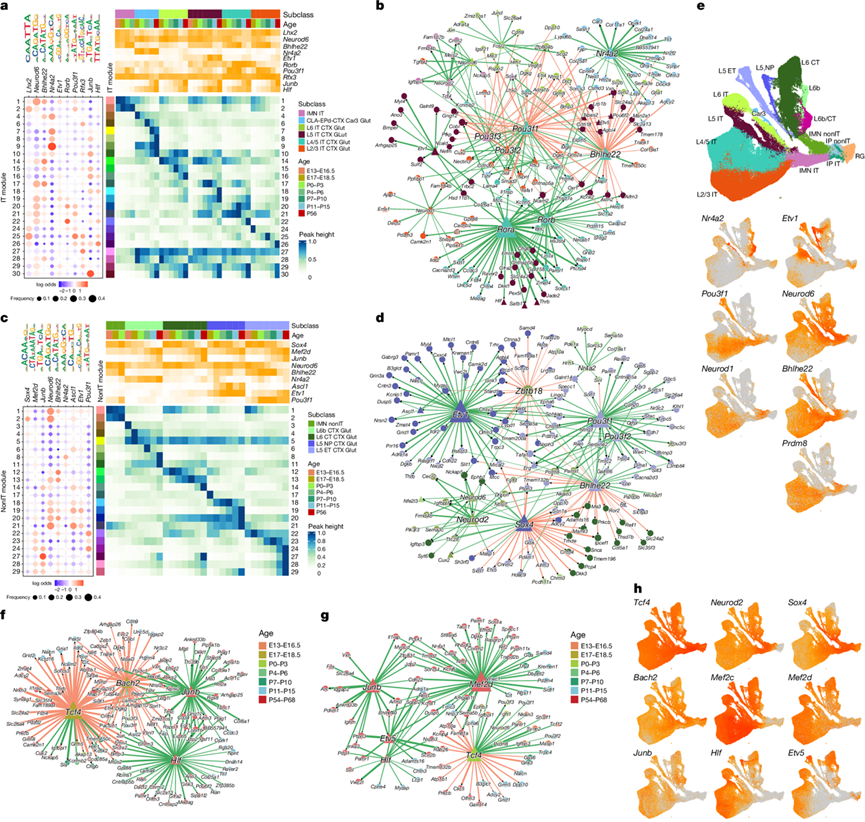
图7. IT 和非 IT 谷氨酸能神经元类型的 GRN。
(a, c) 在IT亚类和非IT亚类中,染色质可及性峰模块中TF DNA结合基序的富集情况。(b, d) IT亚类和非IT亚类的基因调控网络(GRN)。(e) IT 亚类和非 IT 亚类之间共有的细胞类型转录因子调控因子的表达。(f-g) IT 亚类和非 IT 亚类中参与时间调控的GRN。(h) IT 亚类和非 IT 亚类之间共有的时间转录因子调控因子的表达。
+ + + + + + + + + + +
结 论
本研究构建了一个全面且高分辨率的小鼠视觉皮层发育转录组和表观基因组细胞类型图谱,这些数据集涵盖了胚胎期和出生后发育阶段(从胚胎第11.5天到出生后第56天)。本研究鉴定了特定发育阶段新细胞类型出现的分支点以及细胞多样化的分子特征。轨迹图显示,胚胎期的神经发生、胶质细胞发生和早期有丝分裂后成熟以交错平行的方式产生所有细胞类别和几乎所有亚类。在出生后的分化过程中,细胞类型不断精细化,包括在睁眼期和关键期开始时许多细胞类型的后期出现,这表明皮层发育的不同阶段存在持续的细胞类型多样化。在整个发育过程中,特定细胞类型的基因表达和染色质可及性存在协同的动态变化。他们识别出细胞类型特异性且时间分辨的基因调控网络,这些网络通过可及的染色质基序连接转录因子和下游靶基因。
+ + + + +



