

 English
English文献解读|Nat Commun(14.3):蛋白质组学分析揭示预测染色体不稳定性的非小细胞肺癌亚型和肿瘤微环境
✦ +
+
论文ID
原名:Proteogenomic analysis reveals non-small cell lung cancer subtypes predicting chromosome instability, and tumor microenvironment
译名:蛋白质组学分析揭示预测染色体不稳定性的非小细胞肺癌亚型和肿瘤微环境
期刊:Nature Communications
影响因子:14.3
发表时间:2024.11.23
DOI号:10.1038/s41467-024-54434-4
背 景
肺癌是全球主要的健康问题,占全球癌症相关死亡人数的 18%。目前,手术切除(无论是否联合术后辅助治疗)是局部晚期或早期肺癌的一线治疗方法。在晚期病例中,基于基因组学的靶向治疗和使用免疫检查点抑制剂的免疫疗法提供了有希望的治疗选择,通常作为化疗的补充。虽然通过适当的手术和药物治疗,肺癌相关死亡率在持续下降,但局部性、区域性和转移性非小细胞肺癌 (NSCLC) 的 5 年生存率仍然不令人满意(分别为 64%、37% 和 8%),主要是由于治疗后复发。因此,根据分子特征对患者进行分层以制定个性化治疗方案以改善患者生存率越来越受到关注。
实验设计
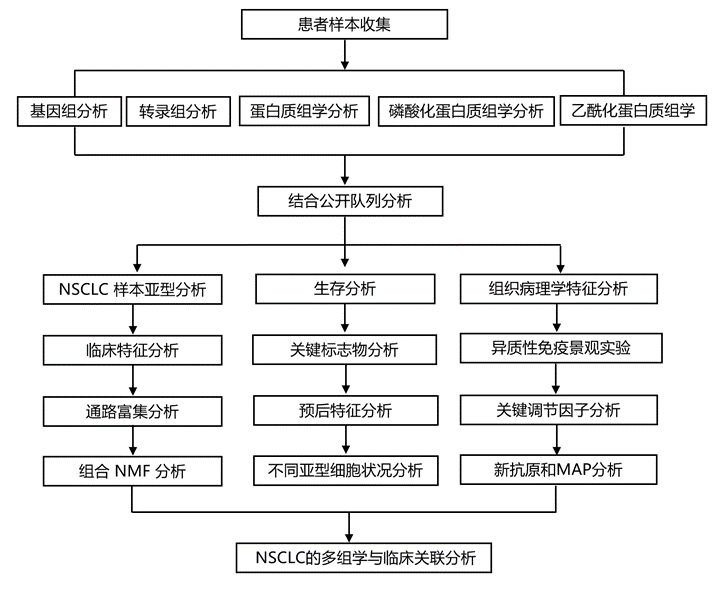
结 果
01

通过多组学方法识别 NSCLC 患者的亚型
研究团队采用了韩国首尔峨山医疗中心确诊为 NSCLC 的 229 名韩国患者的回顾性队列。2010 年至 2019 年期间通过手术采集了肿瘤样本以及匹配的正常组织或血液样本。患者人口统计学、疾病参数(包括组织学和肿瘤淋巴结转移 [TNM] 分期)、生存率和治疗反应表明疾病严重程度和患者人群的覆盖率良好。从组织学上看,肿瘤样本包括 139 个腺癌(LUAD,61%)、63 个鳞状细胞肺癌(LSCC,27%)和 27 个其他类型肿瘤(12%)(图 1a)。 LUAD 病例在性别之间几乎平均分布(66 例男性和 74 例女性),而 LSCC 病例主要见于男性(97%,n = 61),这之前已在其他韩国 LSCC 队列中得到证实。自我报告的吸烟状况表明 61% 的患者有吸烟史,男性和 LSCC 患者的吸烟率更高。基于 TNM 的分期范围从 IA1 至 IVA,大约 40% 的患者患有晚期疾病(IIIA,n = 75;IIIB,n = 9;IVA,n = 11)(图 1a)。大约一半的患者 在手术后病理诊断时有淋巴结转移。 48% 的患者根据 NCCN 指南接受了辅助治疗,包括化疗 (CTx) 或放射治疗 (RTx)。在接受辅助治疗的患者中,54% 的患者出现肿瘤复发,不同治疗方法和不同组织学诊断之间的复发率相似(图 1a)。
他们从样本中生成了基因组、转录组、蛋白质组、磷酸化蛋白质组和乙酰化蛋白质组数据集。通过对 228 个正常邻近组织 (NAT) 匹配的肿瘤和一个仅肿瘤样本进行全外显子组测序 (WES) 生成基因组数据集。他们在 229 个肿瘤中观察到 33301 个包含单核苷酸变异 (SNV) 和插入缺失的体细胞小突变(平均每个样本 145 个)、470836 个拷贝数变异,包括扩增、增益、杂合缺失和纯合缺失(平均每个样本 2056 个)和 2.7 的肿瘤突变负荷 (TMB) 评分。对于转录组分析,他们对 205 个肿瘤和 85 个匹配的 NAT 进行了RNA-seq,以实现深度覆盖(每个样本约 120 M 个读数),从而实现基因表达定量和可变剪接异构体的发现。获得了 60688 个转录本,并根据样本中低计数基因选择了 20088 个转录本进行后续分析。对于使用基于串联质谱标签 (TMT) 的同质异位素标记进行蛋白质组学分析,他们从 229 个肿瘤样本和 26 个匹配的 NAT 收集了蛋白质组学数据。在至少 30% 的样本中观察到总共 10788 个蛋白质、40738 个磷酸位点和 5975 个乙酰化位点,并以与共同参考 (CR) 样本的 log 2比率进行量化(图 1b)。
对于多组学分析,他们整合了蛋白质组学、磷酸化蛋白质组学和乙酰化蛋白质组学数据,并进行了非负矩阵分解 (NMF) 聚类,以识别 229 例 NSCLC 样本中的多组学亚型。他们确定了五种多组学亚型:代谢型(亚型 1)、肺泡样型(亚型 2)、增生型(亚型 3)、缺氧型(亚型 4)和免疫原性(亚型 5)(图 1c),根据基因突变、临床表型和分子通路进行表征(图 1d-e)。
亚型 1 主要由患有EGFR和TP53突变的LUAD 女性(64%,35/55)组成,并且全基因组加倍(WGD)事件的频率很高(即获得一半以上的染色体的现象)(图 1c),表明这是一种染色体不稳定的共同驱动亚型。亚型 1 中CDKN2A拷贝数丢失的显著富集也支持这一观察结果(图 1d)。亚型 2 主要由患有 LUAD(71%,32/45)的患者组成,这些患者患有EGFR突变(49%,22/45)且无 WGD 事件(图 1c),表明这是一种染色体稳定的致癌基因驱动亚型。这些样品显示出明显较低的TP53突变频率(6%)且肿瘤突变负担 (TMB) 比代表其他亚型的样品低得多(图 1d)。尽管两种亚型中EGFR通路均上调,但分子通路在每个聚类中都特异性地富集。亚型 1 表现出参与氧化磷酸化、线粒体基质和细胞呼吸的蛋白质显著上调,表明与代谢通路有关。相反,亚型 2 的特征是 IL -33和Notch通路的磷酸化显著上调(图 1e),表明这些分子特征与早期肿瘤发生和早期疾病有关。
亚型 3 与 WGD 事件和PIK3CA突变显著相关,并且在 LSCC 患者、男性和吸烟者中更为普遍(图 1d)。亚型 3 表现出高度增殖表型,证据是细胞周期相关通路显著富集,包括E2F/MYC靶点、G2M 检查点和细胞周期蛋白依赖性激酶(CDK)靶点通路(图 1e)。因此,该亚型可定义为染色体不稳定的肿瘤抑制基因缺陷型增殖亚型。
亚型 4 与任何特定的 NSCLC 组织学类型无关,但在转移中显著富集。他们发现该亚型中的磷酸化位点在缺氧、PI3K-Akt和中性粒细胞脱颗粒通路中上调,并且蛋白质也在中性粒细胞脱颗粒中富集,表明在促进肿瘤转移中肿瘤迁移、侵袭和代谢方面可能发挥作用。因此,亚型 4 可视为染色体稳定的间充质亚型。
亚型 5 显示肿瘤浸润淋巴细胞 (TIL) 相关模式比例显著升高(图 1d)且免疫相关通路富集, 如 TNFα 通过 NF-κB 信号传导(图 1e), 表明该亚型是高免疫和染色体稳定的肿瘤抑制因子驱动的炎症亚型。KRAS突变在亚型 5 中显著富集, 其中 4 例同时出现TP53突变。STK11和KEAP1突变分别在 1 例中存在,但趋势并不具有统计学意义。
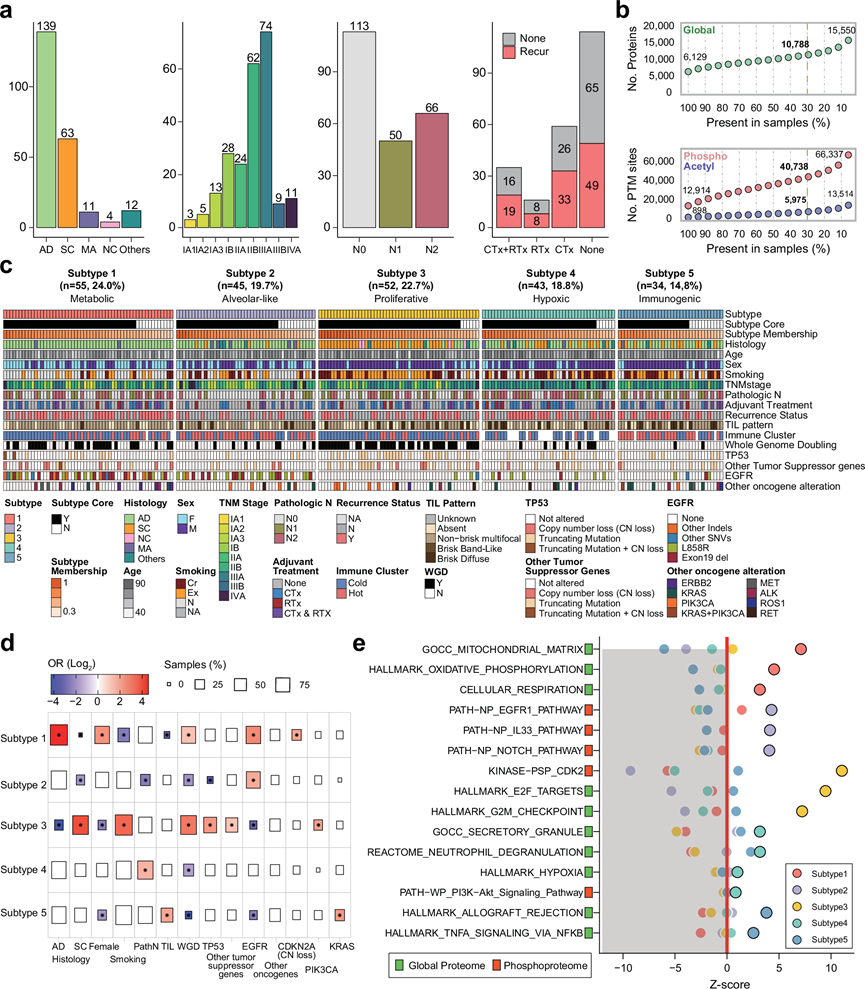
图1. 韩国 NSCLC 患者的多组学亚型识别。
(a) 韩国 NSCLC 患者队列的临床信息摘要。 (b) 在蛋白质组学、磷酸化蛋白质组学和乙酰化蛋白质组学分析中确定的量化蛋白质和 PTM 位点的数量。(c) NMF聚类概述。其他肿瘤抑制基因包括CDKN2A、STK11、KEAP1、RB1、PPP2R1A 和 SMARCA4 。(d) 已识别的五个 NMF 聚类的临床变量富集。(e) 使用 GSVA 和 PTM- SEA对五种亚型进行通路富集分析。
02
一种与预后不良和频繁转移相关的 NSCLC 亚型
他们分析了公开数据中获得的 462 例 NSCLC 患者的多组学数据,并比较了本研究队列与其他研究的 NMF 亚型的主要特征,发现存在显著重叠:腺癌的终末呼吸单元亚型(亚型 1)、炎症亚型(亚型 2 和 5)以及 LUAD 和 LSCC 的增生性亚型(亚型 3)(图 2a)。相比之下,亚型 4 则有所不同,显示出与 EMT 富集 (LSCC)亚型相关的磷酸化特征和来自 LUAD 和 LSCC 炎症亚型的乙酰化特征的富集。他们还通过将研究中的蛋白质组、磷酸化蛋白质组和乙酰化组数据集与先前研究中的数据集整合在一起,对总共447名 NSCLC 患者进行了组合 NMF 分析(以下称为“组合 CPTAC 数据集”)。确定了五种亚型,称为“组合 NMF”(图 2b)。组合 NMF 与特征重叠分析显示出高度一致的模式(图 2a),并证实了四种亚型(亚型 1、2、3、5)与之前确定的 NMF 亚型保持一致(图 2b)。在组合 NMF 分析中,亚型 4 未与 LUAD 或 LSCC 的任何单一亚型聚集(图 2b)。不包括韩国队列(“CPTAC NSCLC”)的比较表明,在包括韩国队列时,77% 的患者保持其亚型分类(图 2c)。此外,亚型 4 始终占韩国 NSCLC 队列的 18.8% (43/229),占无组织学类型的综合 CPTAC 数据集的 18.3% (40/218)。总之,研究结果表明亚型 4 代表需要进一步临床和分子表征的 NSCLC 亚型。
为了检查亚型 4 的分子特征,他们在 NMF 聚类分析中提取了亚型的不同特征(蛋白质、磷酸化或乙酰化)。发现亚型 4 中存在的大多数 NMF 特征是磷酸化位点,表明磷酸激酶相互作用是主要特征。因此,他们使用磷酸蛋白质组学数据研究了此亚型的激酶活性。与其他亚型相比,亚型 4 中两种激酶CSNK2A1和GSK3B显著富集,已知它们可磷酸化PI3K-AKT信号通路中的各种蛋白质(图 2d)。在评估重要激酶活性与表达水平之间的关系(P < 0.05)时,他们观察到中等相关性。基于特征表达的生存分析显示,大多数不利的预后因素是亚型 4 中差异表达的磷酸化位点(91%,104/114)(补充图 2f)。值得注意的是,STE20样丝氨酸/苏氨酸蛋白激酶[SLK (S347)]是一种由CSNK2A1磷酸化的蛋白质,在亚型 4 中显著上调并与不良预后特征相关(图 2e-f)。在组合的 CPTAC 数据集中,他们还发现亚型 4 中SLK (S347) 的磷酸化增加和较差的生存结果(图 2e-f)。SLK介导ErbB2和PI3K通路下游的细胞凋亡并以CSNK2A1依赖的方式激活。最近的研究报告显示,SLK高表达与HER2阳性患者和胶质瘤 的总体生存率降低有关。结合本研究队列和 CPTAC 队列数据以分析 ROC 曲线,发现SLK(S347)不仅在本研究队列中是区分亚型 4 的有效标志物,而且在 CPTAC 队列中也更有效。总之,这些结果表明SLK参与肿瘤进展,可能是亚型 4 特有的关键标志物。此外,亚型 4 显示富含亮氨酸重复序列的不能飞行相互作用蛋白 1 在丝氨酸 581[LRRFIP1(S581)]磷酸化显著上调,这与不良预后特征相关(图 2e-f)。类似地,在组合的 CPTAC 数据集中,在亚型 4 中观察到 LRRFIP1 (S581) 磷酸化升高,这与较差的生存结果相一致(图2e-f)。值得注意的是,LRRFIP1通过调节 Wnt/β-catenin 信号通路来刺激上皮-间质转化 (EMT) 通路。最近的一项研究表明,高LRRFIP1表达与胶质瘤总体生存率降低有关。这些结果共同表明,LRRFIP1也可能有助于细胞侵袭和转移,并可作为亚型 4 特有的关键标志物。
亚型 4 包括HIF-1和PI3K-AKT信号通路中的许多预后特征(图 2g)。在这些不利的特征中,已知显著上调的ANGPT2蛋白在缺氧环境中会增加,这可以通过HIF-1积累促进促血管生成细胞因子(如VEGF)的释放。他们还观察到 K65 处TIMP3乙酰化增加,这可能与ANGPT2诱发的高血压反应有关。相比之下,CDKN1B显著下调,在本研究队列和组合 CPTAC 数据集中是一个有利的预后特征。鉴于CDKN1B的缺失会增加癌症转移的风险, CDKN1B表达降低意味着亚型 4 的晚期表型。
他们根据亚型 4 患者的生存率评估了该亚型的临床意义。韩国 NSCLC 队列中的亚型 4 的生存率明显低于其他亚型(图 2h),可能受分期分布的影响。由于亚型 4 在五种亚型中转移率最高,因此转移可能对不良生存结果产生重要影响。有趣的是,亚型 4 有无转移患者的生存率没有差异(图 2i),这表明转移可能不是导致生存率较差的唯一机制。在比较五种亚型非转移性患者的生存率时,他们发现亚型 4 患者的生存率明显处于劣势(图 2j),表明其他生物学因素可能导致该亚型预后不良。相反,他们在合并的 CPTAC 队列中未发现亚型 4 与其他亚型之间的总体生存率有任何显著差异,这表明种族和治疗记录的差异存在影响。
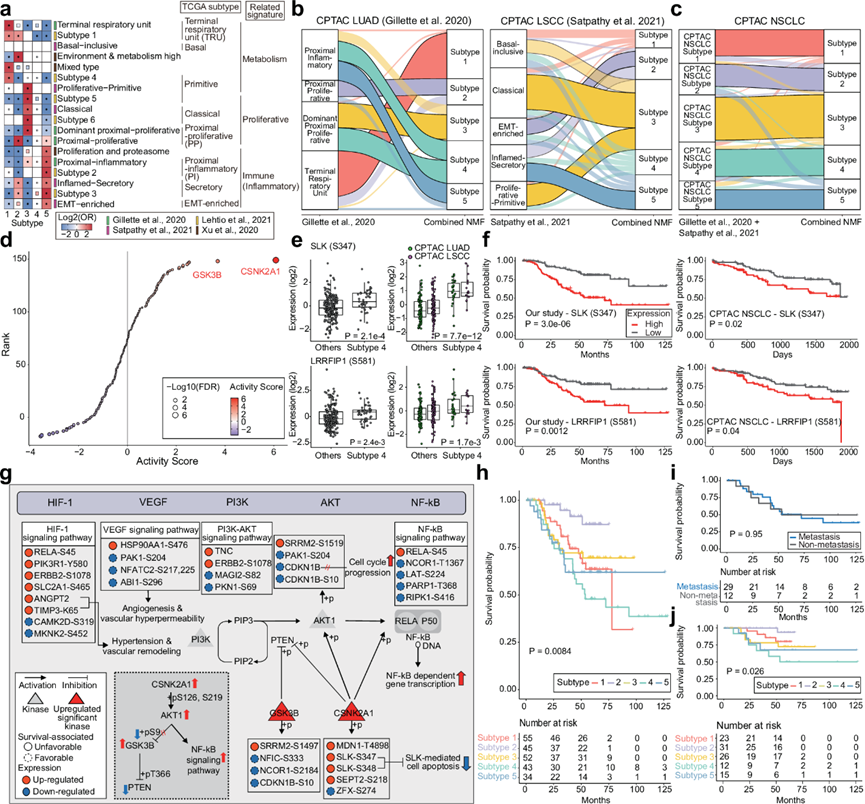
图2. 新型 NSCLC 亚型与较差的生存率相关。
(a) 本研究中的五种 NMF 亚型与先前 NSCLC 多组学研究中确定的亚型之间的亚型特征重叠。(b-c) 根据本研究组合的NMF亚型对样本进行重新分类。(d) 根据磷酸化蛋白质组学数据和激酶-底物网络数据库 (PHONEMeS) 估计的亚型 4 特异性激酶活性分数。(e) LK (S347) 和LRRFIP1 (S581)上含有磷酸化位点的不良预后标志物的表达。(f) 根据不良预后标志物的表达和 CPTAC 数据集(与 LUAD 和 LSCC 整合)得出的癌症特异性总生存期。(g) 不良预后的细胞内信号通路。(h) 亚型之间的癌症特异性总体生存长度表明生存概率(y 轴)随时间(x 轴)发生显著变化。(i-j) 每个亚型中无转移的患者的生存曲线。
03
NSCLC 五种亚型的细胞状况
探索肿瘤微环境对于了解癌症进展的潜在机制和制定有效的治疗策略(不仅针对癌细胞而且针对周围的微环境)至关重要。他们结合了 NSCLC 患者的整合单细胞转录组分析(scRNA-seq)数据集 ,通过对亚型特异性基因与一系列细胞类型特异性基因的比较分析,评估了基于细胞类型特异性的五种 NSCLC 亚型的肿瘤微环境。首先,他们计算了与 NAT 样本相比的每个亚型的差异表达基因(DEG)。正如预期的那样,他们发现所有五种亚型的 DEG 集在肿瘤细胞类型中均显著上调,突出了亚型的整体致瘤程度。
接下来,他们通过比较所有亚型(DEG亚型,例如亚型 1 与其他亚型)来确定每个亚型的 DEG。在亚型 1 和 3 中,DEG亚型在与肿瘤相关的细胞类型中富集(FDR < 0.01,排列):亚型 1 对应 LUAD 和 LUAD 有丝分裂细胞类型,亚型 3 对应 LSCC 和 LSCC 有丝分裂细胞类型,表明肿瘤细胞成分正在活跃增殖(图 3a)。相反,亚型 2、4 和 5 的DEG亚型集在中性粒细胞中富集。虽然这些亚型中存在大量 NAT 相关中性粒细胞 (NAN),但肿瘤相关中性粒细胞 (TAN) 细胞类型在亚型 4 和 5 中富集,其中亚型 4 中所占比例最大。此外,亚型 2 的 DEG亚型组似乎富含肺泡型成纤维细胞和内皮细胞,这意味着这些肿瘤具有与正常肺泡间质相似的基质成分。大多数免疫相关细胞类型在亚型 5 的 DEG亚型组中富集,反映了高比例的 TIL 模式。
他们将这些发现与亚型的组织病理学特征进行了关联(图 3b-m)。亚型 1 和 2 主要是高至中等分化腺癌,其中腺泡和乳头状模式占主导地位(图 3c-d)。LUAD 病例的组织学模式比较表明,鳞状模式在亚型 2 中更为广泛(图 3e)。相反,与亚型 2 相比 ,亚型 1 具有更高比例的高级别组织学模式,包括实性、微乳头状、筛状和复杂的腺状模式(图3f)。与这些观察结果一致,亚型 2 肿瘤表现出较少的淋巴管侵犯(图 3g)和淋巴结转移(图 3h)。亚型 3 主要由鳞状细胞癌和一部分实性为主的腺癌组成(图 3c-d)。亚型 3 肿瘤具有增大的多形性细胞核,有丝分裂活动高,肿瘤坏死频繁(图 3i),这与 DEG亚型分析一致。与亚型 4 和 5 相反,亚型 3 肿瘤的基质成分相对稀少(图 3b-j)。亚型 4 肿瘤具有中等至大量的促纤维化基质成分和不同数量的炎症细胞,而亚型 5 肿瘤在大约一半的病例中表现出免疫细胞成分的高肿瘤浸润(图 3k)。同样,在DEG亚型分析中,亚型4和5的肿瘤更可能伴有中度至高度间质中性粒细胞浸润,但此类肿瘤的比例并不高(图 3l)。总的来说,他们获得了五种亚型的代表性组织学图像,反映了它们的组织病理学特征和临床相关性。
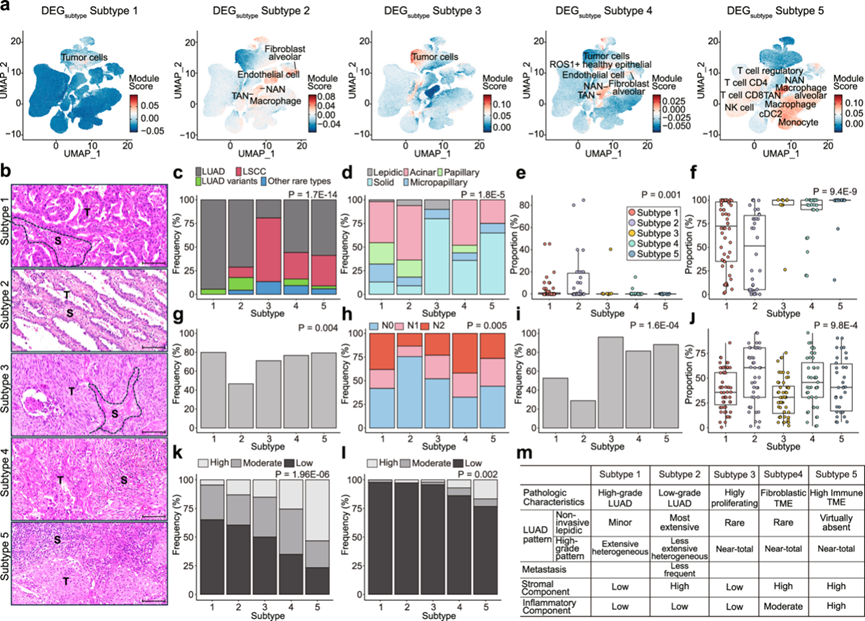
图3. 细胞类型特异性亚型特征图。
(a) 使用每个 DEG亚型对亚型 1-5 特有的单细胞类型进行 UMAP 图。(b) 代表性组织学图像。(c) 每种亚型中具有不同病理诊断的样本的比例。(d-f) 每种亚型中 LUAD 的组织学模式。(g-i) 淋巴管侵犯、淋巴结转移和肿瘤坏死在亚型 2 肿瘤中较少见,这也意味着亚型 2 肿瘤处于临床早期、无进展阶段。(j) 显微镜下,亚型 2、4 和 5 肿瘤中的基质成分更为广泛。(k-l) 肿瘤浸润淋巴细胞和基质中性粒细胞浸润在亚型 5 肿瘤中最为广泛。(m) NSCLC亚型的组织病理学特征总结。
04
NSCLC 亚型全基因组加倍的蛋白质组学特征
多组学分析将亚型 3 确定为以 WGD 事件为特征的增殖亚型。在亚型 3 中,75% 的患者 (39/52) 表现出 WGD(图 4a)。拷贝数特征分析显示亚型 3 与 CN14 特征之间存在显著关联(图 4b),这表明在单次基因组加倍事件之前,存在高倍性、相对较大的片段大小(> 40 Mb)和整个染色体或染色体臂规模的丢失。具体来说,与其他亚型相比 ,亚型 3 在染色体 3q 上表现出扩增,并且SOX2扩增和TP53突变同时出现显著富集(图4c-d)。此外,在整合了 CPTAC LUAD 和 LSCC 数据集的组合 NMF 分析中,亚型 3 也显示出高频率的 WGD(图 4a)和 CN10(28%,18/64)和 CN16 特征(17%,11/64)的富集,这分别表明在单基因组和两倍基因组加倍之前存在局部和染色体丢失。在本研究和先前的研究中,亚型 3 的 TMB 均高于其他亚型。
与之前的研究一致,亚型 3 显示细胞周期途径中的蛋白质、PTM 和激酶上调(图 4ef)。参与此亚型的关键蛋白质和 PTM 包括SRSF1(蛋白质 S199 和 K179)、SRSF2(T25 和 S26)和XPO1(蛋白质 K693)。SRSF1和SRSF2都是原癌基因,它们调节肿瘤抑制基因、激酶和激酶受体的剪接模式,使其成为致癌异构体,从而促进细胞周期活动。在此亚型中, XPO1在蛋白质和乙酰化蛋白质水平上均上调(图 4g)。在许多恶性肿瘤中都观察到XPO1的过度表达,抑制它可能会减少细胞增殖并促进增殖亚型的细胞周期停滞。为了验证这一假设,他们从 SG Medical, Inc.(韩国首尔)的类器官生物库中选择了由 WGD 阳性 LUAD 和 LSCC 患者以及 WGD 阴性对照产生的肿瘤类器官。在用靶向 XPO1 抑制剂 selinexor 处理类器官后,发现 WGD 阳性 LSCC 类器官的药物敏感性高于其他样本(图 4h)。这一发现表明,用 selinexor 靶向XPO1可能是一种有前景的 LSCC 和 WGD 肿瘤患者治疗方法,这值得进一步研究其潜在的临床应用。
在较小程度上,亚型 1 还包括患有 WGD 的患者(图 4a)。亚型 1 在 CN15 标记(图 4b)中显著富集,倍性分布均匀(绝对拷贝数从 2 到 9 以上),片段大小为 1 到 10 Mb。该亚型主要见于 LUAD 患者,与EGFR和肿瘤抑制基因(TP53或CDKN2A)的共病突变有关(图 4d),这意味着亚型 1 在 LUAD 中普遍存在,并且 WGD 是通过 LUAD 特异性肿瘤演变事件激活的。在组合 NMF 分析中,他们验证了 WGD 阳性患者的比例(图 4a) 以及先前 LUAD 和 LSCC 研究中亚型 1 中 WGD 相关特征 CN16 的富集。
全基因组缺失表征了两种主要的 NSCLC 亚型,但每种亚型似乎都有不同的驱动基因和潜在通路(图 4i)。亚型 1 的特点是EGFR的帧内缺失和拷贝数增加率较高,且肿瘤抑制基因的突变负担较高。亚型 1 中显著上调的激酶包括RPS6KA3 (它介导由EGFR激活启动的细胞生长信号传导)、CDK1、CDK2 和 CDK4,表明细胞周期加速。此外,与 DNA 修复和自噬分别相关的CDK12和PIK3C3也显著上调。由于细胞增殖增加会导致缺氧和营养不足36,这些激酶可能在 WGD 反应中发生激活,以促进细胞适应营养缺乏的环境并稳定基因组,最终导致细胞存活。同样,与DNA 损伤和营养缺乏反应相关的激酶与CDK1、CDK2 和 CDK4一起在亚型 3 LSCC 样本中发生激活。然而,亚型 3 中 WGD 的假定驱动突变是TP53突变和 3q 染色体上的拷贝数扩增,许多细胞周期基因位于此处,包括SOX2、ATR、STAG1、GSK3B、TFDP2 和 MCM2。由于 LUSC 肿瘤往往在 3q 臂中具有拷贝数增益,亚型 3 可能受益于 WGD 后的 3q 增益,因此具有比亚型 1 样本更高的 WGD 分数。此外,与亚型 1 不同,CHEK1 和 EEF2K在经历 WGD 后发挥重要作用。已知EEF2K在饮食限制下通过磷酸化使EEF2失活,从而有助于细胞存活。总体而言,这些结果表明亚型 1 和 3 分别代表在 LUAD 和 LSCC 肿瘤中经历 WGD 的增殖亚型,并且 selinexor 可能是治疗亚型 3 患者的有效方法,这需要进一步的验证研究。
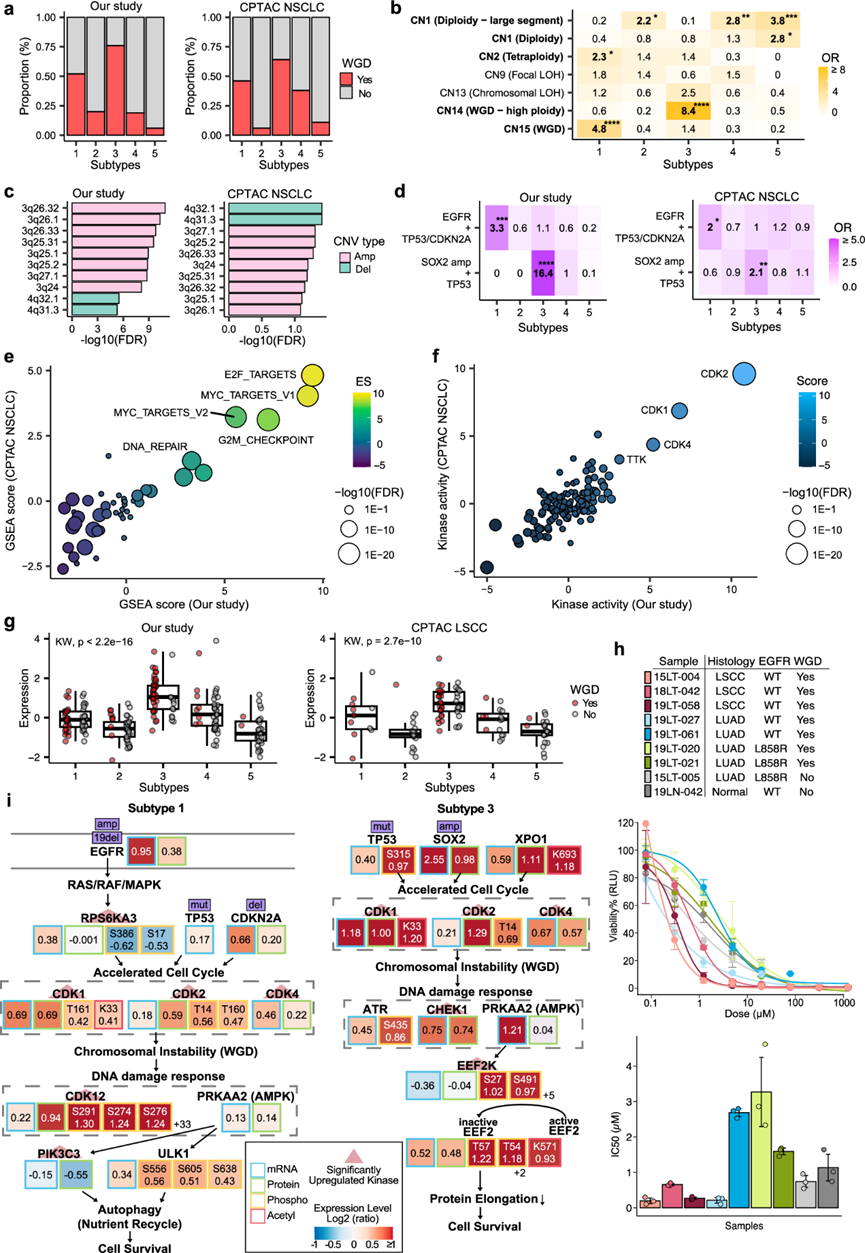
图4. NSCLC 亚型全基因组加倍 (WGD) 的蛋白质组学特征。
(a) 条形图显示本研究和 CPTAC NSCLC 患者中每种多组学亚型的 WGD 分数。(b) 五种多组学亚型的拷贝数特征重叠。(c) 亚型 3 中前 10 个最显著富集的拷贝数变异(CNV)。(d) 不同队列中每种亚型中富集的共突变。(e) 基因集富集分析 (GSEA) 揭示了两个队列的亚型 3 中的上调和下调通路。(f) 特异性激酶活性得分。(g) XPO1 蛋白表达升高。(h) 肺类器官的样本信息(上)、药物反应曲线(中间)和 selinexor(XPO1抑制剂)的 IC50(下),突出显示了 WGD 阳性 LSCC 类器官的更高灵敏度。(i) 亚型 1 LUAD 肿瘤和亚型 3 LSCC 肿瘤背后的WGD相关通路。
05
NSCLC 中的异质性免疫景观
了解肿瘤免疫微环境 (TIME) 对于癌症亚型的分子表征至关重要。为了分析 NSCLC 患者的 TIME,进行了两项基于细胞类型和通路的聚类分析,并从精选的基因特征推断出富集分数。他们在 205 个肿瘤和 85 个 NAT 中确定了三个主要免疫聚类,即热肿瘤富集 (HTE)、冷肿瘤富集 (CTE) 和 NAT 富集(图 5a)。基于细胞类型和基于通路的聚类之间的 HTE 和 CTE 免疫聚类有相当大的重叠。与 CTE 肿瘤相比,HTE 肿瘤在细胞类型水平上主要富集于 CD8+ 和 CD4+ T 细胞、调节性 T (Treg) 细胞、B 细胞、自然杀伤 (NK) 细胞、中性粒细胞、树突状细胞 (DC)、单核细胞和巨噬细胞。在基于通路的聚类结果中,HTE 聚类显示出比 CTE 聚类更强的免疫相关通路激活。由于细胞类型富集分数是根据精选的基因特征推断出来的,他们使用来自多个 NSCLC 研究的 scRNA-seq 数据来确认免疫聚类的细胞类型特异性。从 HTE (DEG HTE) 和 CTE (DEG CTE) 聚类中的前 300 个 DEG 中,他们研究了每个 DEG 在 scRNA-seq 数据中的富集情况。 DEG HTE集富集于 CD8+ 和 CD4+ T 细胞、调节性 T 细胞、B 细胞、NK 细胞、中性粒细胞、DC、单核细胞和巨噬细胞,而 DEG CTE集主要富集于上皮细胞,包括 LUAD 和 LSCC 肿瘤细胞(图 5b)。此外,从基因特征推断出的每种细胞类型的 DEG 在 scRNA-seq 数据中的相应细胞类型中富集。细胞类型富集评分也与 TIL 水平高度相关(图 5b)。这些结果表明单细胞和免疫组织化学分析根据细胞类型评分实现了良好的免疫聚类。
为了分析 NSCLC 患者免疫景观的预后价值,对每个免疫细胞富集评分以及免疫聚类进行了总体和无复发患者生存率的测试(图 5c)。HTE 状态是患者生存的最有利因素,CD8+ 和 CD4+ T 细胞、B 细胞和 NK 细胞的高富集与良好预后呈正相关,而上皮细胞或中性粒细胞的富集则呈负相关。所有基于免疫相关通路的富集评分均与患者生存期无显著相关性,但大多数显示出与延长生存期正相关的相关性。已知 CD8+ 和 CD4+ T 细胞及免疫相关通路高富集的 HTE 肿瘤与良好的预后相关。此外,先前的研究报告称,Treg 在 HTE 肿瘤中富集,但具有免疫抑制作用且与预后不良有关。他们发现 HTE 肿瘤中 Treg 富集的趋势一致(图 5a-b);Treg 富集的 HTE 患者的生存率比 Treg 水平低的患者更差。
通过分析各自的调节因子,可以更好地阐明 HTE 和 CTE 免疫聚类之间的差异。为了确定患者免疫景观的假定调节因子,他们分析了 RNA 和蛋白质表达数据,并根据蛋白质在相关靶标转录中的系统性影响推断蛋白质调控活性。对基因表达或活性以及细胞类型或免疫聚类的富集分数进行了回归分析。他们发现大多数免疫调节因子(主要包括癌细胞配体和免疫细胞受体)与 HTE 肿瘤和大多数免疫细胞呈正相关,但与上皮细胞无相关。值得注意的是, VEGF-A的表达和活性与 HTE 肿瘤呈负相关,这与观察到的其他免疫调节因子的模式形成对比。VEGF -A与 TIL 呈负相关,对癌症风险有潜在影响。具体而言,10 种免疫调节因子与免疫聚类状态表现出显著相关性,不仅在本研究队列中,而且在其他肺癌多组学研究中也呈现出一致的相关方向(图 5d)。
他们分析了免疫调节因子与已知驱动突变之间的关联,以确定免疫调节因子变化的假定机制。由于亚型 5 具有免疫原性,他们仅考虑最活跃或最受抑制的免疫调节因子,以确定将其与其他亚型区分开来的潜在调节机制。最终,这三种免疫调节因子与八个驱动基因之一的突变显示出显著的相关性,在其他多组学 NSCLC 队列中也观察到了一致的模式(图 5e)。在这些免疫调节因子中,SMARCA4突变与SLAMF7在 RNA 和蛋白质水平上的表达呈正相关,并且在本研究队列和独立队列中都与 HTE 状态呈正相关(图 5f)。根据这些结果,SMARCA4是免疫调节因子SLAMF7的潜在调节因子,与 HTE 状态相关。
他们进一步分析了多组学亚型之间的免疫景观分布(图 5g)。HTE 特征基因(DEG HTE)的表达在具有免疫原性特征的亚型 5 中最高。该亚型还显示出最高的 CD8+ 和 CD4 + T 细胞、Treg 细胞、B 细胞、NK 细胞、中性粒细胞和巨噬细胞标志基因活化。中性粒细胞和 Treg 分别与所有患者和特别是 THE 患者的预后不良相关(图 5c),在亚型 5 中也发现富集。尽管亚型 5 表现出最强的 THE 肿瘤信号,但这种富集可能与在此亚型中观察到的中等生存特征有关。此外,在亚型 5 中观察到了最高的SMARCA4突变和SLAMF7过表达频率(图 5g)。在将本研究队列与最近的多组学研究的队列相结合的综合队列中,免疫聚类内细胞类型的分布及其与多组学亚型的相关性是一致的。结果表明,HTE 肿瘤与 NSCLC 患者的预后较好呈正相关。这些肿瘤富含免疫原性亚型 5,这也与假定的调节因子SMARCA4的突变呈正相关,SMARCA4 靶向免疫调节因子SLAMF7。

图5. 不同 NSCLC 亚型和群体的免疫聚类和细胞类型概况。
(a) 基于细胞类型和通路富集评分的免疫亚型。对205个肿瘤和85个肿瘤旁正常 (NAT) 样本进行基于细胞类型的聚类。(b) UMAP 图。(c) 基于细胞类型的免疫聚类中各种细胞类型相关的总生存期和无复发生存期的风险比。(d) 免疫调节因子的 RNA 表达、蛋白质表达和蛋白质活性与免疫聚类状态的相关性。(e) 免疫调节因子的表达或活性与驱动突变状态之间的相关性。(f) 左:箱线图显示 野生型或突变型SMARCA4样本中SLAMF7的 RNA 和蛋白质表达;右:箱线图显示HTE 和冷肿瘤富集 (CTE) 样本中SLAMF7的 RNA 和蛋白质表达。 (g) (左)箱线图和气球图显示多组学亚型中 HTE 和 10 种细胞类型的标记基因的平均表达。(右)条形图和箱线图分别显示多组学亚型中SMARCA4的突变频率和SLAMF7的 RNA/蛋白质表达。
06
新抗原和免疫聚类的多组学分析
新抗原是由肿瘤细胞产生的肿瘤特异性抗原,是影响癌症患者免疫状况的关键因素48。因此,他们使用多组学数据集预测了NSCLC 队列中的新抗原。使用 WES 数据预测源自编码区体细胞突变的新抗原,并通过 MS 分析进行确认,以确定它们是否表达为“确认的新抗原候选物”的肽。此外,他们还鉴定了源自非编码和未注释转录本的肽,这些肽不同于典型的新抗原。其中,他们选择了“隐蔽性 MHC I 类相关肽 (MAP)”,在之前的研究中将其为预测与 MHC I 类分子结合的非典型肽。根据之前确定的表达阈值鉴定了“确认的隐蔽性 MAP”。他们推断出 85430 个新抗原候选物和 775 个隐蔽性 MAP,并根据匹配的转录本注释了隐蔽性MAP 的来源。未注释的异构体、假基因和非翻译区(UTR)隐蔽性MAP 来源的 90.97%.
他们测试了新抗原候选物、已证实的新抗原候选物、隐蔽性 MAP 和已证实的隐蔽性 MAP 与患者生存率的关联(图 6a)。有趣的是,只有已证实的隐蔽性 MAP 与生存率提高呈现出强烈的正相关性,虽然患者的隐蔽性MAP 数量明显较低,但是他们还在三名以上的患者中发现了 12 个已证实的隐蔽性 MAP,称为复发性隐蔽性 MAP,其中一些来自同一来源基因。此外,复发性隐蔽性 MAP 的存在与延长生存期显著相关(图 6b),表明隐蔽性 MAP 在 NSCLC 患者中的预后价值。
他们还研究了隐蔽性 MAP、免疫聚类和细胞类型富集之间的关联。隐蔽 MAP 与大多数免疫细胞类型(包括 HTE 状态)呈正相关,尽管有些相关性较弱或不显著(图 6c)。在本研究队列中,根据免疫聚类标准和隐蔽 性MAP 负荷分层后评估患者预后。隐蔽性 MAP 负荷和 HTE 状态高的患者生存期最长,而隐蔽性 MAP 负荷和 CTE 状态低的患者生存期最差(图 6d)。
他们通过评估基于隐蔽性 MAP 负荷和免疫聚类的综合状态的多组学亚型的分布来确定综合免疫景观与多组学亚型之间的关联。多组学亚型 5 在低隐蔽 MAP 负荷和 HTE 组中最富集(图 6e)。有趣的是,当用抗原加工、呈递机制 (APM) 和 TIL 模式取代免疫聚类时,获得了相似的生存趋势和多组学亚型分布,这是评估新抗原免疫激活潜力的关键因素。值得注意的是,表现出中等生存率的亚型 5 患者具有低隐蔽 MAP 负荷、HTE 和激活的 APM。这与单细胞和组织病理学分析中观察到的大量 TIL 比例一致(图 3a-k)。亚型 5 患者除了 APM 外,还通过 NF-κB 显示出 TNF-α 通路的高活性。因此,虽然具有 HTE 状态的活跃免疫景观,但与该亚型相关的中等生存率似乎与低负荷隐蔽 MAP 和高富集免疫抑制性 Treg 和中性粒细胞有关(图 6f)。值得注意的是,使用辅助化疗或放化疗可显著提高亚型 5 肿瘤患者的生存率(图 6g)。相反,在接受辅助化疗或放化疗的其他亚型患者中,生存率没有显著改善。尽管统计意义很小,但亚型 5 在接受辅助化疗的总体人群中也表现出与其他亚型相比最有利的预后。这强调了亚型 5 可能与辅助化疗的临床益处有关。这些结果暗示,了解患者预后和多组学亚型需要对 TIME 进行多因素考虑。此外,结合预测前后的 HLA 等位基因分布与之前关于韩国人群或肺癌研究的报告一致,从而证实了本研究结果的可靠性。
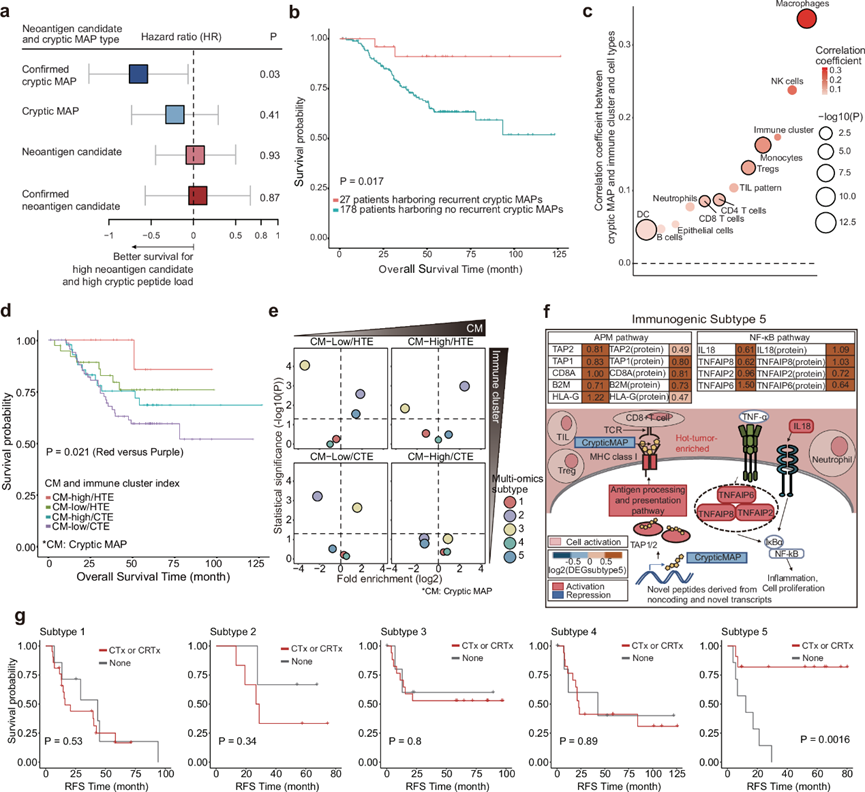
图6. 新抗原和隐蔽性 MAP的临床相关性及其与多组学亚型的关联。
(a) 生存期分析。 (b) Kaplan-Meier 曲线显示两组患者的生存率。(c) 隐蔽性 MAP 数量与 10 种细胞类型和免疫聚类的富集分数之间的相关性。(d) Kaplan -Meier 曲线显示按隐蔽性 MAP和免疫聚类分层的四组患者的生存模式。(e) 针对多组学亚型的富集分析。(f) 患者的特征。(g) 根据多组学亚型分类的患者治疗状态得出的无复发生存率的 Kaplan-Meier 曲线。
+ + + + + + + + + + +
结 论
本研究对来自韩国 NSCLC 队列的 229 名患者进行了全面的多组学分析,并确定了五种富含 WGD、致癌基因、转移和免疫微环境的 NSCLC 亚型。磷酸化蛋白质组数据集对亚型识别最有帮助,而整体蛋白质组和乙酰化组数据也在解码已识别亚型的信号通路方面发挥了关键作用。对免疫微环境的全面表征揭示了不同分子亚型的各种免疫细胞组成和新抗原负荷,这预测了不同的预后。免疫亚型表现出热肿瘤富集状态和更高的辅助治疗功效。
+ + + + +



